|
Происхождение спирулы
|
|
| heyfetzed | Дата: Четверг, 24.12.2009, 23:54 | Сообщение # 1 |
|
Группа: Проверенные
Сообщений: 21
Репутация: 0
Статус: Offline
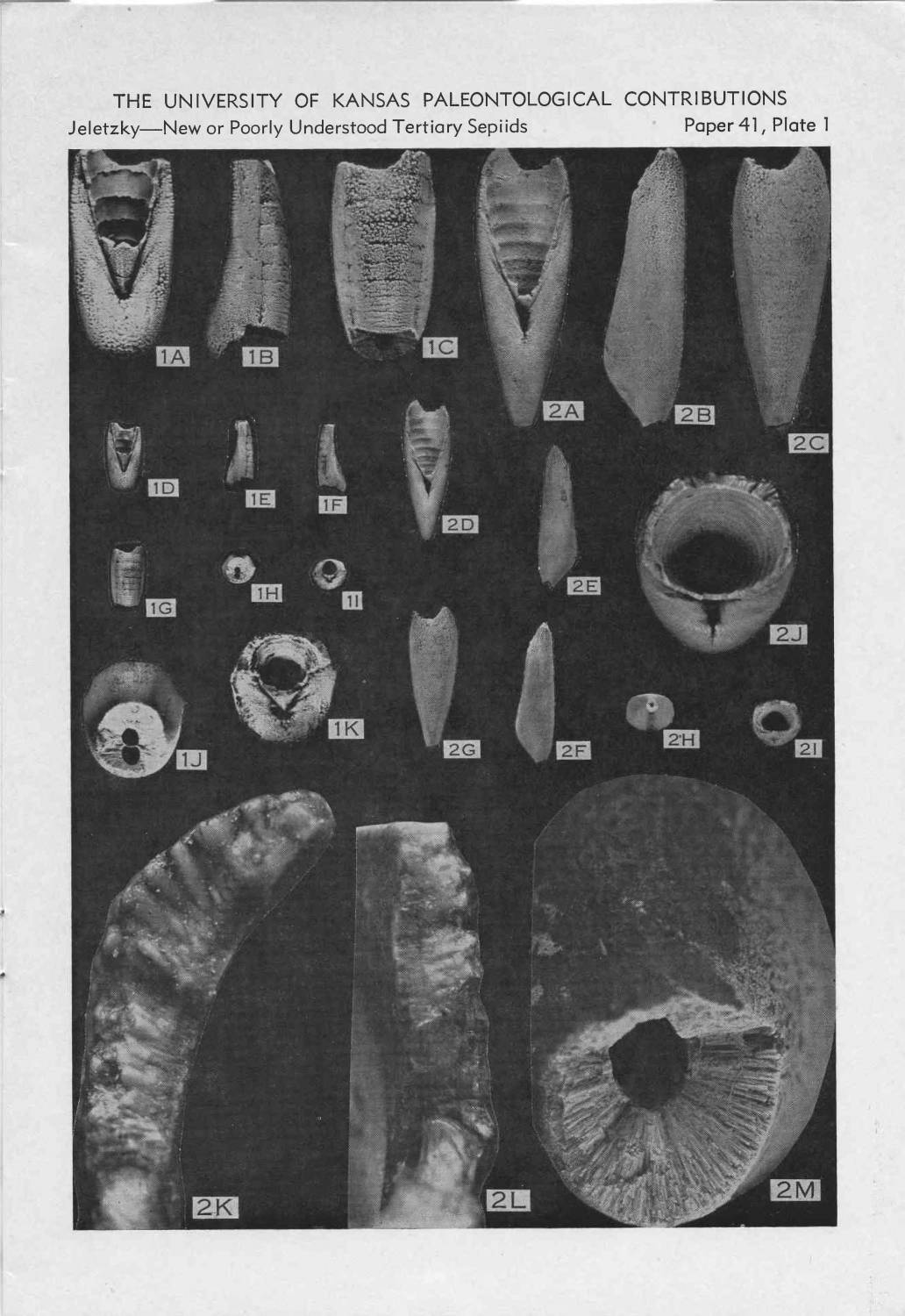
| Э. О. Хейфец Происхождение спирулы Нормальной тенденцией в эволюции моллюсков с внутренней раковиной является ее редукция вплоть до полного исчезновения. У головоногих такая тенденция на первых порах осложняется появлением внутреннераковинных придатков, приспосабливающих раковину-поплавок к новому положению. Соответственно, мы расписываем уравнение, дабы упростить его. Кажущимся исключением представляются Belemnoseina, белемнитоподобные организмы с изогнутой камерной частью раковины (фрагмоконом). Считается, что изгиб прогрессирует, и что наиболее продвинутым видом является спирула — современная форма с внутренней спиральной раковиной. В отличие от белемнитов, являющихся руководящими ископаемыми, белемнозеины крайне редки [1], т. к. их рострум состоит не из кальцита, а из менее стойкого арагонита. Еще меньше шансов сохраниться в ископаемом состоянии у хрупкой раковинки спирулы.
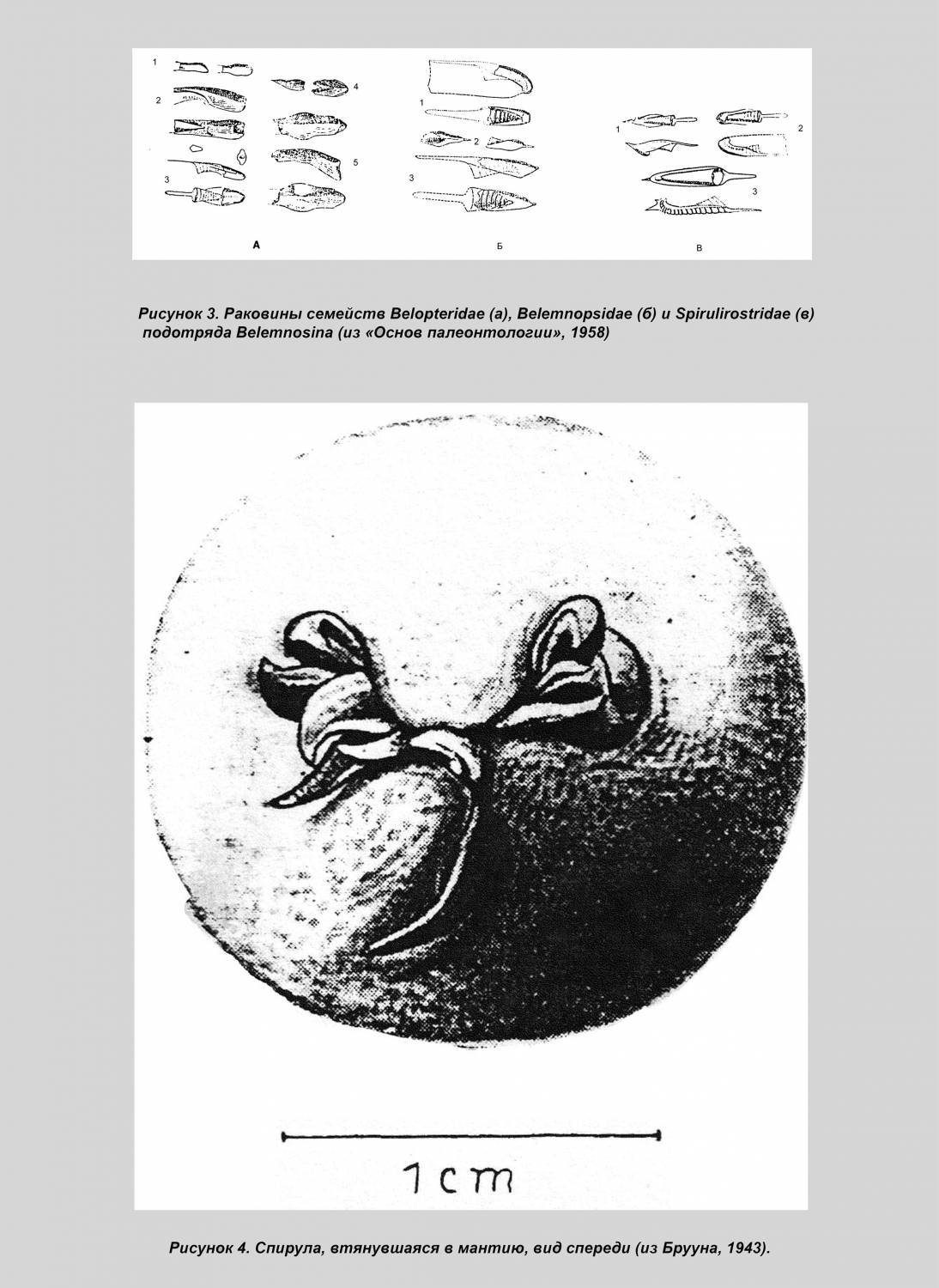
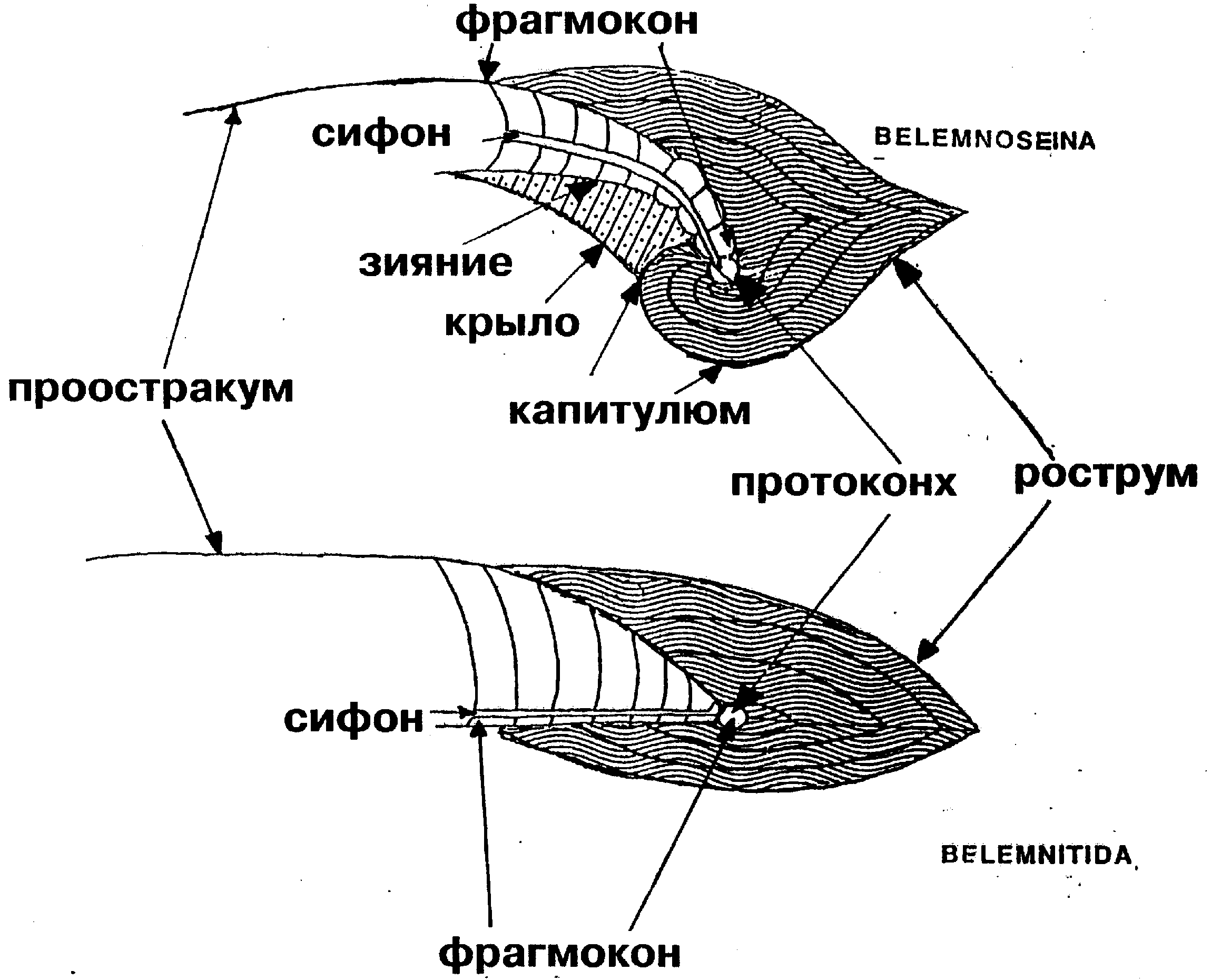
Такая парадоксальная тенденция как спирализация внутренней раковины, утратившей защитную функцию, уникальна и заслуживает объяснения. Тем более требует теоретического обоснования вывод, сделанный на основании малого фактического материала. Однако он лишь постулируется на основании ископаемых находок. Истинным мотивом явилось, скорее всего, явное сходство белемнозеин и белемнитид, от которых белемнозеин первоначально и производили [1], а также построение монофилетического древа колеоидей. Против принятой версии свидетельствует морфология раковины белемнозеин. Так их рострум, несмотря на крайнюю изменчивость, не редуцируется, тогда как смена изогнутости более консервативной камерной части — фрагмокона — налицо (рис. 2).
В отличие от белемнитов рострум белемнозеин, формировавшийся натянутым на раковину мантийным мешком, не развивается на вогнутой брюшной части фрагмокона (рис. 3), подобно тому, как тетива не прилегает к вогнутой части лука. Эта особенность характерна и для белемнозеин с выпрямленной раковиной и их потомков, настоящих каракатиц (Sepiidae), где изгиб фрагмокона не может воспрепятствовать развитию рострума. Т. о., выпрямлённые формы происходят от более изогнутых, а не наоборот. Такая тенденция обычна у внутреннераковинных моллюсков. Впрочем, брюшная борозда на роструме белемнитов может свидетельствовать об их происхождении от форм с зиянием, вызванным изгибом фрагмокона. Примечательно также отсутствие у спирулы характерных внутринераковинных придатков: рострума — даже в остаточном виде; в форме боковых крыльев и наплыва на наружной стороне, как у значительно более редуцированной раковины настоящих каракатиц (сепиона), равно, как и отсутствие проостракума, фиксирующего раковину в коже спины, и потому небесполезного для данного организма. Судя по всему, отсутствие внутреннераковинных придатков у спирулы первично. Спирула оказывается не потомком белемнозенин, а представителем предковой группы, происходящей от наружнораковинных спиральных форм. К таковым относятся аммониты, для которых, как и для других потомков бактритоидей характерна зародышевая камера (протоконх), отличающая их от прочих спиральных головоногих. С наружнораковинными моллюсками спирулу сближает целый ряд особенностей. Так, её внутренняя раковина не вписывается в контуры тела, а выступает со спиной и брюшной стороны тела. Т. о. её диаметр вместе с покрывающей её кожицей больше такового туловища.
В случае опасности спирула втягивает голову с конечностями в мантию, которая закрывается мантийными клапанами [2] (рис. 4).
Здесь уместно вспомнить классическую реконструкцию Траута (Trauth), согласно которой аммониты закрывали устье раковины не головной лопастью, как наутилиды, а мантийным оперкулюмом, цельным (анаптих) либо в виде створок (аптихи). В пользу такого вывода свидетельствует удлинённая жилая камера аммонитов. При хищном жизни это соответствовало развившимся щупальцам. Во втянутом состоянии голова оказывалась в глубине раковины и не могла закрыть её устья.
Такую реконструкцию отверг Шиндевольф на том лишь основании, что мантия не может одновременно синтезировать арагонитовую раковину и кальцитовый оперкулюм. Новая версия была принята, несмотря на слабость аргумента — синтез арагонитового фрагмокона и кальцитового рострума хорошо известен у белемнитов. Аптихи и анаптихи были интерпретированы как челюсти аммонитов [3: 138], что противоречит находкам у тех же экземпляров челюстей, типичных для головоногих [4: 182]. Впоследствии возникают книжные версии о происхождении оперкулюмов аммонитов непосредственно из челюстей [5]. Спирула служит дополнительным аргументом в пользу классической версии, а последняя — подтверждением аммонитного происхождения спирулы.
Спирула является единственным современным представителем подкласса аммонитов, который считался вымершим в конце мезозоя. Более того, данный подкласс не представляет собой слепой ветви, заканчивающейся спирулой. К последней близки каракатицы, осьминоги, а также закрытоглазые кальмары, в отличие от настоящих или открытоглазых кальмаров (Оеgорsіdа), которые могут быть потомками белемнитоподобных форм. Если последние происходят непосредственно от прямых бактритоидей, происхождение внутреннераковинных головоногих явно полифилетично. Тем не менее, как было сказано выше, брюшная борозда на роструме белемнитов может свидетельствовать о происхождении от спиральных форм. В 1982 г. Райтнер и Энгезер предположили, происхождение спирулид от аммонитов, в отличие от прочих белемнитоподобных. Свой вывод авторы обосновывают тем, что у спирулид и аммонитов протоконх открыт, тогда как у белемноморф он отделён от прочих камер вторичной мембраной [7]. Т. о. спирулиды не могли произойти от специализированных белемноморф. Отсюда, однако не следует, что спирулиды происходят от аммонитов. Открытый протоконх является исходным для всей бактритоидной ветви, в т. ч. и для прямораковинных бактритоидей, а также предков высокоспециализированных белемноморф.
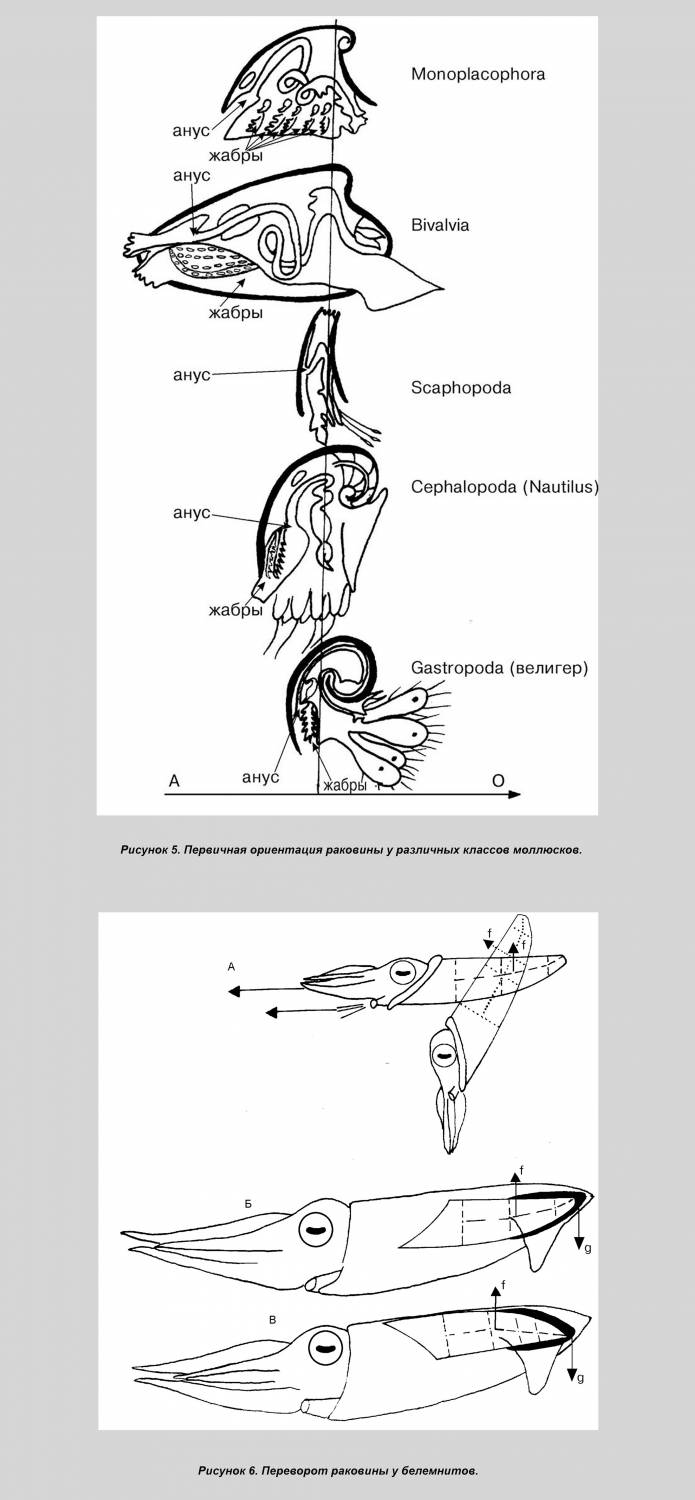
Примечательно, что очевидное сходство спирулы с аммонитами не обсуждается. Более того, спирулиды произведены от прямораковинного гренландиабеллитеса. Т. о., вывод о родстве спирулы с аммонитами вносится контрабандой. Такая тактика позволяет избежать полемики с оппонентами по ключевым вопросам и, к сожалению, становится весьма обычной в современном теоретизировании. В отличие от наружнораковинных наутилид и аммонитов, завиток спирулы является не экзо-, а эндогастричным, т. е., закрученным не на спину, а на брюхо. Такая ориентация может служить аргументом в пользу происхождения от белемнитоподобных форм (при принятии за их непосредственных наружнораковинных предков прямых бактритоидей), т. к. спиной край лучше развит у всех прямо- внутреннераковинных головоногих, в т. ч. у современных каракатиц и кальмаров. Тем не менее, раковина белемнитов также реориентирована. У их наружнораковинных предков — бактритоидей брюшной край был развит сильнее, что, в общем, соответствует первичной ориентации раковины у моллюсков, обнаруженной мною. Такая ориентация определяется положением жабр, абсорбирующих соли, идущие на постройку раковины и выделяемые через анус вследствие несовершенной выделительной системы (рис. 5). Поскольку у внутреннераковинных головоногих устье сильно уменьшено, брюшной край приближен к спинному, а жабры отдалены от обоих, метаболизм не препятствовал реориентации раковины, которая в любом случае имела место, и которую надлежит объяснить. Проще объясняется реориентация у белемнитов в случае их происхождения от прямых наружнораковинных форм (в противном случае, данная версия будет упрощенной, но достаточно наглядной).
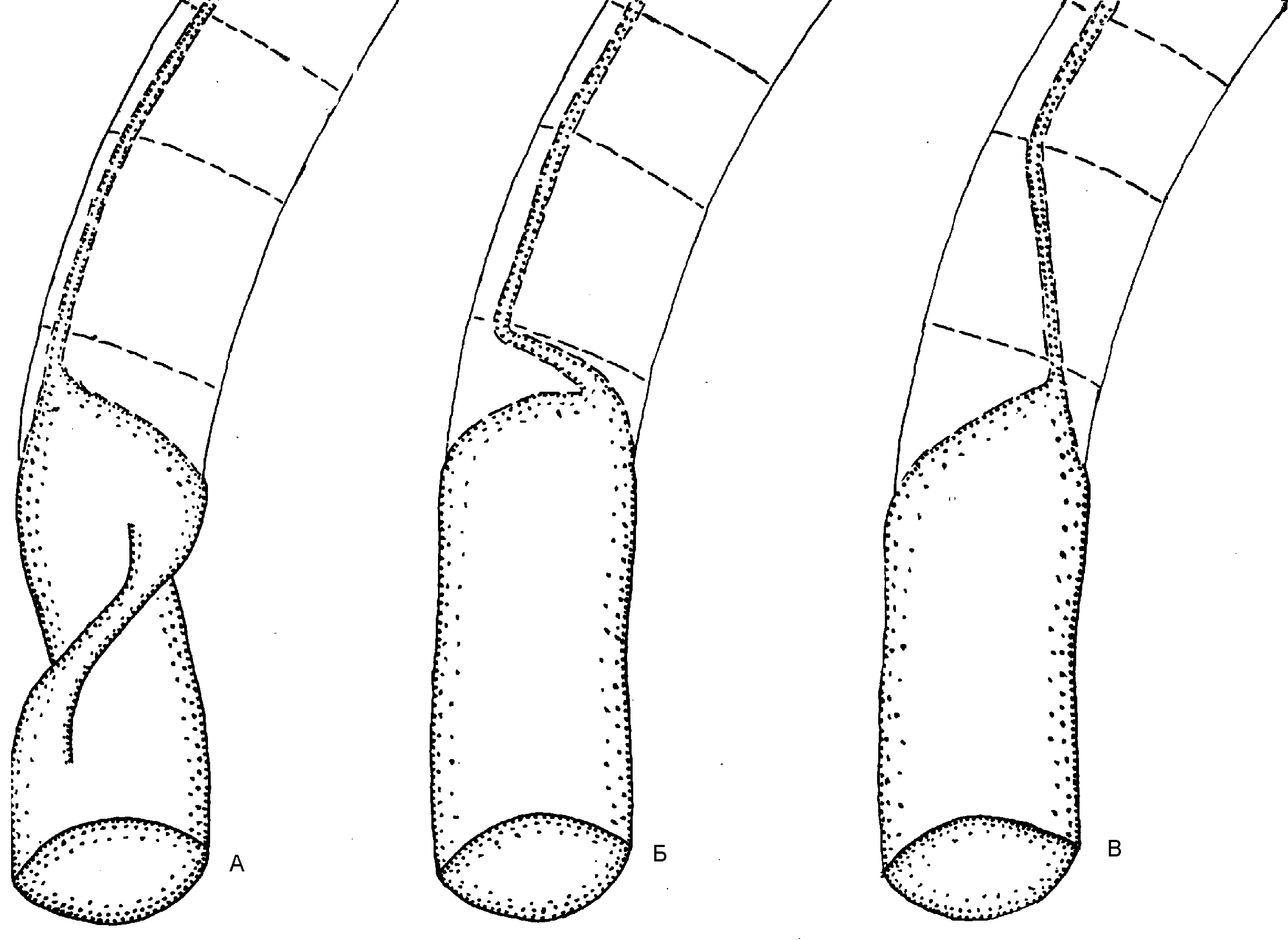
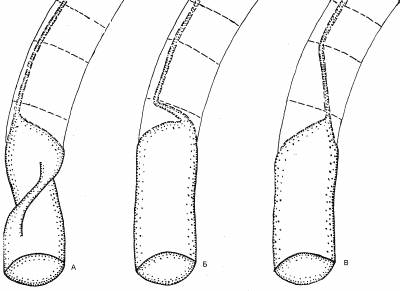
Само наличие рострума, отклонявшего раковину в горизонтальное положение, уже свидетельствует о перераспределении сил плавучести. У наружнораковинных предков белемнитов раковина принимала горизонтальную позицию лишь во время броска и сохраняла её благодаря импульсу. Плавники, а затем и рострум сделали такую позицию постоянной. В результате более развитая и потому выпуклая брюшная часть погрузилась вниз вместе с центром плавучести. Стремясь подняться, он разворачивал раковину (рис. 6). Этому благоприятствет и то обстоятельство, что раковина у внутреннераковинных лежит свободно в мантийном мешке, не прирастая к её стенкам [6: 33].
Несколько сложнее реориентация происходила у спиральных форм: Постоянному погружению раковины под мантию предшествовало временное её обволакивание, уменьшавшее трение о воду и защищавшее раковину от обрастаний. Животное стало быстрее передвигаться и, одновременно, медленнее втягиваться в раковину. Уменьшение защитной функции раковины дополнительно стимулировало скоростное движение. Тело перестало помещаться в раковине, которая сдвинулась к аборальному концу. В новой позиции центр плавучести оказался позади центра тяжести, отклоняя тело от горизонтали. Более равномерное распределение плавучести было достигнуто раскручиванием спирали (геликоническая раковина) (рис. 7, А).
Кроме того, если у наружнораковинных аммонитов большой диаметр раковины располагался вертикально, то у внутреннераковинных он сместился к оси тела (как ныне у спирулы). Произошло это из-за увеличения диаметра раковины вследствие её раскручивания и дополнительно способствовало равномерному распределению плавучести. В результате поднятия устья раковины центр плавучести опустился. Если у наружнораковинных форм он был расположен поверх устья, то у спирул раковина оказалась облегчённой: тело больше не занимает жилую камеру, а значит, центр плавучести сместился к последнему обороту (рис. 7, А). На рисунке 7, Б показана реориентация спиральной раковины, но переворот, скорее всего, происходил ещё до полного формирования завитка. Образ жизни спирулы. Считается, что в естественных условиях спирула зависает вниз головой в соответствии с распределением плавучести в раковине и не способна к нормальному передвижению. Этому противоречит стайный образ жизни спирулы, а также наличие светящегося органа, явно служащего для дальней коммуникации. Его функция закономерно объявлена загадочной.
Такое представление основывается на непосредственных наблюдениях, проведенных датским учёным А. Брууном в аквариуме. Логично было предположить артефакт: спирулу извлекли с большой глубины, она испытала температурный шок, прошла сквозь слой жалящих кишечнополостных, на что указывал сам автор и что должно было сказаться на наблюдениях. Первоначально я сделал именно такой вывод [8]. Тем не менее, обратившись к источнику год спустя, прочитал: «...При перенесении в аквариум они /особи спирулы/ зависали в вертикальной позиции, раковиной вверх... неподвижные и как будто безжизненные». Это описание и легло в основу существующей версии. Однако вскоре моллюски приходили в себя, и картина менялась: «Как и другие каракатицы, спирула часто производит лёгкие пульсирующие движения, внезапно бросаясь в любом направлении: вверх, вниз или из стороны в сторону... обычно животное двигалось задним концом вперёд... прижав к нему плавники /отсюда, мощность реактивного двигателя (воронки) достаточно велика, чтоб преодолеть сопротивление отклоняющей силы плавучести /. Реже спирула... делала бросок головой вперёд» [2: 4] (см. рис. 8).
По-видимому, пассивная защита спирулы в сочетании с версией о ее происхождении от активных форм привела к выводу о малоподвижном образе ее жизни и, в результате, — к игнорированию непосредственных наблюдений. Верно обратное: в случае активного образа жизни спирулы ее пассивная защита является не новейшей адаптацией, а наследием предков, подобно способности подвижных и кусачих мягкопанцирных черепах втягивать голову в панцирь. Функция светящегося органа. Как было сказано в начале предыдущего раздела, светящийся орган спирулы ранее принимался за присоску. Будучи расположенным на аборальной стороне тела, т. е. максимально удалённым от глаз и щупалец, он явно не служит освещению поля зрения либо приманиванию добычи. Не предназначен он и для защиты от врагов, которых одни головоногие ослепляют, вымётывая светящиеся чернила, а другие дезориентируют свечениям брюха на фоне светлого же неба. Для этих целей видимость светящегося органа слишком ограничена.
Более того, при допущении вертикальной ориентации спирула просто не сумеет увидеть светящийся орган другой особи, поскольку последний направлен вверх, тогда как глаза — вбок. Ненамного улучшает положение и нормальная ориентация: в этом случае спирула сможет увидеть погружённый в присосковидную впадинку светящийся орган другой особи лишь, если их позиции взаимноперпендикулярны, а её глаз находится прямо напротив аборального конца другой особи.
Между тем, спируле необязательно видеть светящийся орган непосредственно. Достаточно сориентироваться по лучику света от него. С другой стороны ориентация хищников (в частности спирулу часто пожирают тунцы) по такому потайному фонарику затруднительна. Так, при приближении хищника спирула втягивается в мантию и мгновенно разворачивается вертикально, выводя светящийся орган из поля зрения хищника.
Сказанное возможно проверить непосредственными наблюдениями с борта подводной лодки.
В статье 8 светящийся орган ошибочно был назван люминофором, тогда как принятым термином является фотофор, на что мне указал доктор биологических наук Анистратенко В. В. (Институт Зоологии им. И. И. Шмальгаузена НАН Украины).
Соответствующие корни происходят от греческого слова «fos» (род. пад. — «fotos») и латинского — «lumen», что в обоих случаях означает «свет». Однако первый корень в большей степени относится к свету, пришедшему извне: фоторецептор, фотосинтез, тогда как второй — к свету, излучённому самим организмом — биолюминесценция. Т. о. термин «фотофор» возник из-за невосприятия этимологии, благодаря чему он способствует путанице и поощряет педантизм, отдающий преимущество выучке перед пониманием. Предлагаю исправить допущенную неточность, и заменить термин «фотофор» на «люминофор». Благодарность Над этой статьёй я начал работать в Израиле, где провёл несколько лет. Существенную помощь оказали сотрудники Иерусалимского Университета: профессор Й. Геллер — в оформлении статьи и доктор Х. Минис, предоставивший важные источники из своей библиотеки. При оформлении иллюстраций меня консультировал художник-анималист В. Фергюссон.
Всем им выражаю свою искреннюю признательность. Библиография 1. Основы палеонтологии. Моллюски — Головоногие, ч. II,/ под ред. Орлова Ю. А. М.; Гостехиздат, 1958. —
359 с.
2. Bruun A. The biology of Spirula spirula. Dana-report, 1943. — 44 p.
3. Lehmann U. The Ammonites. Their life and their forms. Cambridge; Cambridge University Press, 1982. — 466 р.
4. Друщиц В. В., Догужаева Л. А. Аммониты под электронным микроскопом. М.; МГУ, 1981. — 240 с.
5. Adolf Seilacher, Ammonite aptychi: how to transform a jaw into an operculum. American Journal of Science,
Vol. 293-A, 1993, P. 20-32, см. ткж. http://earth.geology.yale.edu/~ajs/1993/11.1993.02Seilacher.pdf (формат
поврежден, файл открывается в HTML).
6. Несис К. Н. Краткий определитель головоногих моллюсков. М.; Лёгкая и пищевая промышленность.
1982. — 353 c.
7. Reithner J., Engeser T., Phylogenetic trends in phragmocone-bearing coleoids (Belemnomorpha). Neues Jahrb.
Geol. Paläont. 164, 1982. – рp. 156 –162
8. Хейфец Е. О. Первісна орієнтація черепашки у молюсків
та її реорієнтація в контексті еволюційної морфології, Вісник Житомирського педагогічного університета
2002 р. № 10, С. 212 –216. Иллюстрации: Возникли сложности. Прикреплю в теч. 24 ч.
Сообщение отредактировал heyfetzed - Пятница, 25.12.2009, 13:22 |
| |
| |
| AlexIpp | Дата: Воскресенье, 27.12.2009, 11:46 | Сообщение # 2 |

Группа: Администраторы
Сообщений: 579
Репутация: 0
Статус: Offline
| Здравствуйте, Эдуард 
Я дам некоторые комментарии к вашей статье, частично - повторяя те, которые приводил вам раньше в переписке: для того, чтобы полноценно поучаствовать в дискуссии могли и другие посетители форума. Это был бы очень интересный опыт обсуждения статьи или идей на Интернет-форуме.
Писать буду небольшими кусками.
Думаю, для начала нашим посетителям будет любопытно посмотреть, на что похожи объекты, которые мы обсуждаем. Это действительно нечасто встречающиеся формы
Итак, третичные ростровые спирулы (табличка из "Основ палеонтологии"):
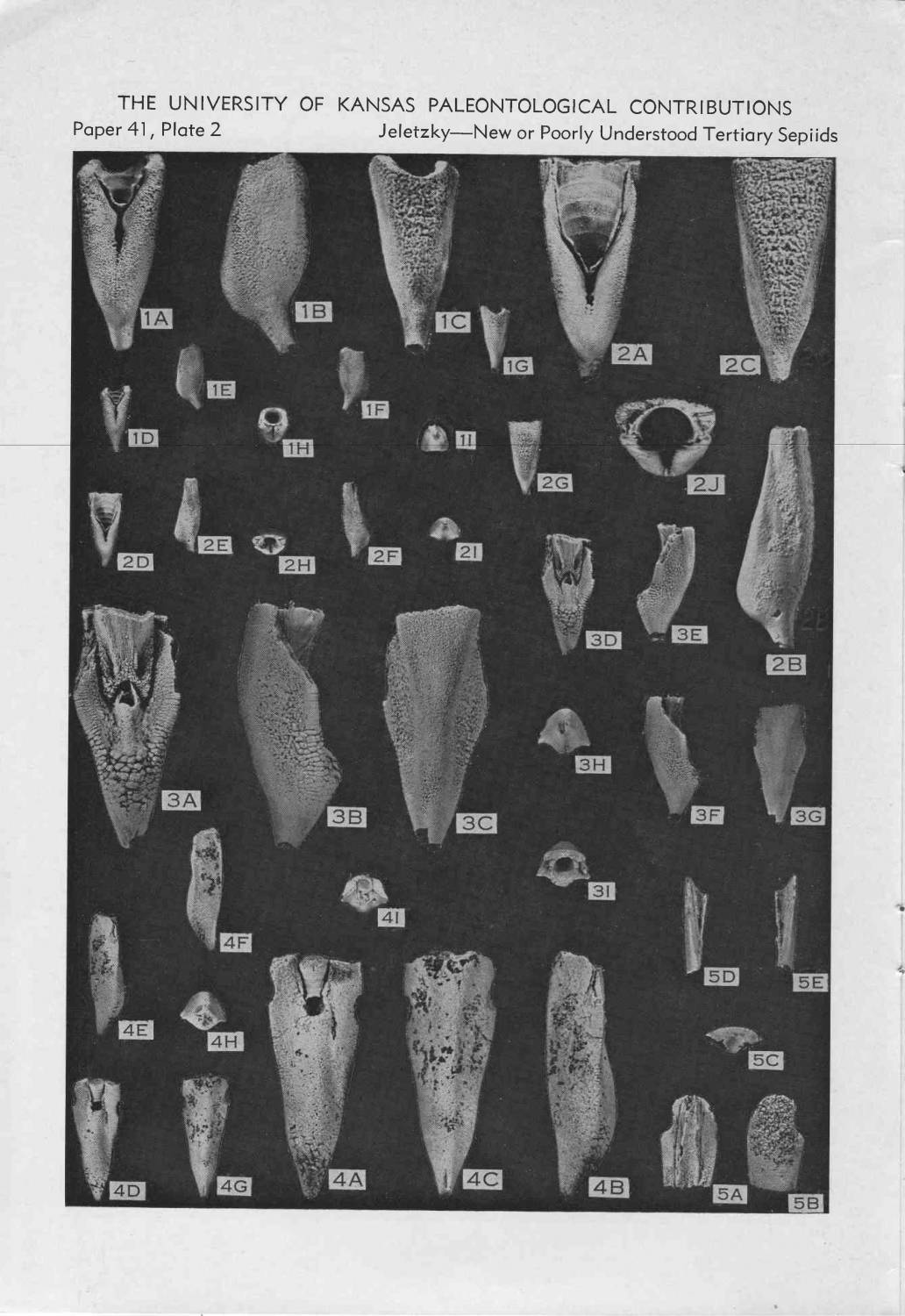
  тоже третичные спирулы - из США (Jeletzky, 1969):
 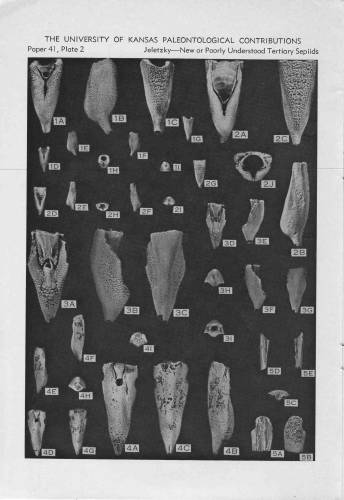
всё о серпулидах & белемнитах
|
| |
| |
| AlexIpp | Дата: Воскресенье, 27.12.2009, 11:55 | Сообщение # 3 |
|
Группа: Администраторы
Сообщений: 579
Репутация: 0
Статус: Offline
| Современная спирула и ее раковины:
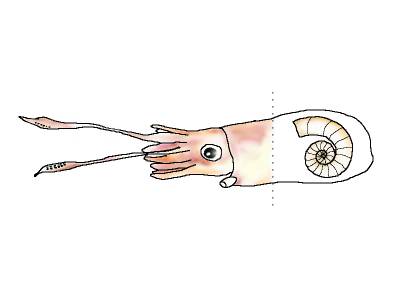 
  ===================================
Общее впечатление о статье.
Мне кажется, тема очень интересная и актуальная, и подана с неожиданной и необычной точки зрения, однако почему-то вы выбрасываете из рассмотрения большое количество как накопленных фактических данных, так и гипотез других исселдователей. А ведь до вас над происхождением спирулид бились лучшие и гениальнейшие головоногие умы XX века - Нэф, Елецкий, и продолжают их дело головы начала XXI века - Бизиков, Догужаева, Фухс и другие... Наконец, я К сожалению, сейчас накопленный научный опыт и багаж знаний все реже позволяет одним взмахом пера перевернуть представления в какой-либо области знаний. Многие выводы указанных людей кажутся хорошо обоснованными, тогда как вы на их работы даже не ссылаетесь и никак не обсуждаете изложенных в них положений.
Я думаю, ваши представления и выводы сильно изменятся после того, как вы детально ознакомитесь с ними.
всё о серпулидах & белемнитах
|
| |
| |
| AlexIpp | Дата: Воскресенье, 27.12.2009, 12:06 | Сообщение # 4 |
|
Группа: Администраторы
Сообщений: 579
Репутация: 0
Статус: Offline
| Quote (heyfetzed) Считается, что изгиб прогрессирует, и что наиболее продвинутым видом является спирула — современная форма с внутренней спиральной раковиной. В отличие от белемнитов, являющихся руководящими ископаемыми, белемнозеины крайне редки [1], т. к. их рострум состоит не из кальцита, а из менее стойкого арагонита.
из третичных отложений описано более 70 (!) номинальных видов ростровых спирулид, относящихся к ~15 родам. Они часто встречаюстя в отдельных осадочных бассейнах (Франция, Англия, СшША) - их изображения регулярно попадаются в каталогах ископаемой фауны XIX века.
Это называется - "крайне редки"? Если речь про бывшее СССР - да, пожалуй.
Quote (heyfetzed) Еще меньше шансов сохраниться в ископаемом состоянии у хрупкой раковинки спирулы.
"хрупкие" раковинки глубоководных спирул претерпевают длительную транспортировку и в больших количествах выбрасываются на пляжи западного побережья Австралии. Кроме того, они найдены в ископаемом состоянии, эти находки неединичны, сделаны в разных регионах мира (Европа, Япония) и во всех случаях их возраст - миоцен, т.е. они моложе ростровых форм. Quote (heyfetzed) Такая парадоксальная тенденция как спирализация внутренней раковины, утратившей защитную функцию,
главная функция внутренней раковины - опорная, плюс это часть гидростатического аппарата. В чем защитная функция? Не понимаю, совсем
Quote (heyfetzed) Против принятой версии свидетельствует морфология раковины белемнозеин. Так их рострум, несмотря на крайнюю изменчивость, не редуцируется, тогда как смена изогнутости более консервативной камерной части — фрагмокона — налицо (рис. 2).
ростр как раз постепенно редуцируется от палеоцена к эоцену и олигоцену Quote (heyfetzed) В отличие от белемнитов рострум белемнозеин, формировавшийся натянутым на раковину мантийным мешком, не развивается на вогнутой брюшной части фрагмокона (рис. 3)
у сприулид постепенно происходит смещение внутренностей (т.е. веса тела) под поплавок (фрагмокон), а противовеса, соотвественно - наверх поплавка. Нижняя его часть теперь окружена не кожистой секретирующей, а нормальной мускульной мантией. Поэтому в рострах снизу образуется зияние.
всё о серпулидах & белемнитах
|
| |
| |
| AlexIpp | Дата: Воскресенье, 27.12.2009, 19:29 | Сообщение # 5 |
|
Группа: Администраторы
Сообщений: 579
Репутация: 0
Статус: Offline
| Quote (heyfetzed) Впрочем, брюшная борозда на роструме белемнитов может свидетельствовать об их происхождении от форм с зиянием, вызванным изгибом фрагмокона.
а как быть с белемнитами, у которых борозда на спинной стороне? А если борозды есть на спинной и брюшной сторонах одновременно? Quote (heyfetzed) отсутствие у спирулы характерных внутринераковинных придатков: рострума — даже в остаточном виде; в форме боковых крыльев и наплыва на наружной стороне, как у значительно более редуцированной раковины настоящих каракатиц (сепиона), равно, как и отсутствие проостракума, фиксирующего раковину в коже спины, и потому небесполезного для данного организма.
функция проостракума - создавать твердо-эластичную продольную опроу тела, что важно в первую очередь для активных пловцов. Спирула таковым не является, поэтому у нее проостракум редуцирован. Кстати, у ископаемых ростровых спирулид проостракумы найдены
Quote (heyfetzed) Спирула оказывается не потомком белемнозенин, а представителем предковой группы, происходящей от наружнораковинных спиральных форм.
однако стратиграфическое распространение ископаемых форм свидетельствует об обратном. Кроме того, спирулиды на сегодня найдены в отложениях вплоть до верхней юры, а некоторые исследователи (Л.А. Догужаева) к спирулидам по микроструктурам относят и некоторые каменноугольные формы. Наиболее древние из известных достоверных спирулид (кимеридж Центральной России) - это ростровые формы с хорошо выраженным проостракумом ну про аптихи mhorn лучше напишет Quote (heyfetzed) Спирула служит дополнительным аргументом в пользу классической версии, а последняя — подтверждением аммонитного происхождения спирулы. Спирула является единственным современным представителем подкласса аммонитов, который считался вымершим в конце мезозоя
ОК. тогда:
1. Где у спирулы аммонитовая линия? У прямых (гетероморфных) аммонитов - бакулитов и др. - аммонитовая линия выражена отлично!
2. Если считать, что эндогастрическая раковина спирулы повернулась в теле на 180 град. в поперечном направлении, то почему вдруг брюшной сифон аммонитов, максимально приближенный к внешнему краю, оказывается у экзогастрической спирулы на внутренней стороне спирали??? Quote (heyfetzed) В 1982 г. Райтнер и Энгезер предположили, происхождение спирулид от аммонитов, в отличие от прочих белемнитоподобных. Свой вывод авторы обосновывают тем, что у спирулид и аммонитов протоконх открыт, тогда как у белемноморф он отделён от прочих камер вторичной мембраной [7]. Т. о. спирулиды не могли произойти от специализированных белемноморф. Отсюда, однако не следует, что спирулиды происходят от аммонитов
протоконх "открыт" и у ростровых третичных спирулид Quote (heyfetzed) спиной край лучше развит у всех прямо- внутреннераковинных головоногих, в т. ч. у современных каракатиц и кальмаров. Тем не менее, раковина белемнитов также реориентирована. У их наружнораковинных предков — бактритоидей брюшной край был развит сильнее, что, в общем, соответствует первичной ориентации раковины у моллюсков, обнаруженной мною. Такая ориентация определяется положением жабр, абсорбирующих соли, идущие на постройку раковины и выделяемые через анус вследствие несовершенной выделительной системы (рис. 5). Поскольку у внутреннераковинных головоногих устье сильно уменьшено, брюшной край приближен к спинному, а жабры отдалены от обоих, метаболизм не препятствовал реориентации раковины, которая в любом случае имела место, и которую надлежит объяснить.
вообще ничего не понял
Quote (heyfetzed) Образ жизни спирулы.
способность спирулы к реактивному движению на коротких промежутках времени никак не противоречит пассивному образу жизни. А ведь большую часть времени спирула именно пассивна. Quote (heyfetzed) Функция светящегося органа
главное - не с какой стороны расположен орган, а что благодаря ему организм в целом имеет некое поле свечения. И видно именно оно, безотносительно к тому, где орган расположен. Кстати, обзор каждого глаза спирулы, судя по форме, составляет почти полную полусферу, то есть два глаза, расположенных на боковых сторонах, должны давать полный обзор окружающего пространства
всё о серпулидах & белемнитах
|
| |
| |
| heyfetzed | Дата: Вторник, 29.12.2009, 17:30 | Сообщение # 6 |
|
Группа: Проверенные
Сообщений: 21
Репутация: 0
Статус: Offline
| По какой-то причине статья не редактируется. С опозданием прикрепляю иллюстрации к "Происхождению спирулы". Надеюсь, разместить по месту назначения. Благодарю Алексея Павловича за превосходные фотографии. На вопросы отвечу позже.
Хейфец Эдуард.
P. s. С наступающим Новым Годом!
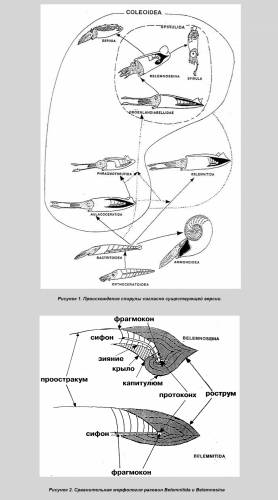  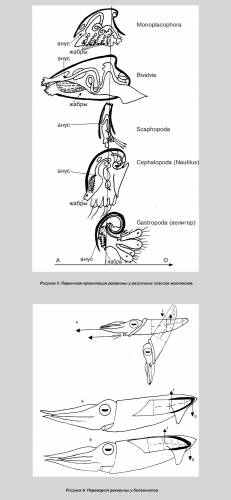 
Сообщение отредактировал heyfetzed - Среда, 30.12.2009, 00:18 |
| |
| |
| heyfetzed | Дата: Вторник, 29.12.2009, 17:31 | Сообщение # 7 |
|
Группа: Проверенные
Сообщений: 21
Репутация: 0
Статус: Offline
| Случайно продублировал сообщение. Без нового текста не удается сохранить.
Сообщение отредактировал heyfetzed - Среда, 30.12.2009, 00:07 |
| |
| |
| heyfetzed | Дата: Пятница, 08.01.2010, 23:27 | Сообщение # 8 |
|
Группа: Проверенные
Сообщений: 21
Репутация: 0
Статус: Offline
| Уважаемые оппоненты!
Прошу прощения за поздний ответ: на Новый Год интернет был забит, и войти я не сумел. Потом был занят на работе.
Я благодарен за Вашу высокую оценку темы, и подхода к ней.
Что до вопросов, то мне представляется затруднительным, в первую очередь, их количество. Ведь надлежащий ответ раза в четыре превысит вопрос — это по длине, плюс еще время на обдумывание. Постараюсь, все же ответить на них, а в дальнейшем предлагаю сосредоточиться на таких моментах, которые способны прояснить ситуацию, а не запутать одного из собеседников.
Что до качества, то судите сами. Начну с замечания об отсутствии ссылок на труды предшественников. Работая над темой, я просмотрел немалое число источников. В обзорной работе им нашлось бы подобающее место. Здесь же вынужден был сосредоточиться на основной теме. Подобающего обоснования происхождения спирулы не нашел. Полагаю, возникло оно стихийно: логично было предположить, что прямораковинные белемниты происходят от прямораковинных же форм. Более редкие белемнозеины и спирула были попросту подчинены сложившемуся представлению. Явное противоречие основным тенденциям эволюции осталось незамеченным.
Т. о. речь может идти лишь об исправлении частной недоработки. Факты же, добытые предшественниками, отрицать не намерен.
N. B. «росчерк пера» длился несколько лет. Так, я не сразу пришел к правильной картине распределения плавучести (вначале не учел ряд мелких факторов), но на ошибки мне не указал ни один из рецензентов — с ними справился сам. Другие обходили основные доводы десятой дорогой. Перехожу к конкретным замечаниям.
По поводу редкости предполагаемых предков спирулы Вы пишете: «из третичных отложений описано более 70 (!) номинальных видов ростровых спирулид, относящихся к ~15 родам. Они часто встречаются в отдельных осадочных бассейнах (Франция, Англия, США) - их изображения регулярно попадаются в каталогах ископаемой фауны XIX века.
Это называется - "крайне редки"? Если речь про бывшее СССР - да, пожалуй. Как палеонтолог, я могу достоверно судить по отсутствию ископаемых спирулид и сепиид в моих сборах (в отличие от многочисленных белемнитов, аммонитов и пр.) об их редкости. Другое дело, отдельные места, не делающие погоды в стратиграфии.
Чем б. СССР хуже прочих мест? Тортон (средний Миоцен) был не менее богат, чем современное Средиземное море. Сошлюсь также на авторитетного и беспристрастного свидетеля — «Основы палеонтологии». Резюмируя состояние изученности группы, автор раздела Г. Я. Крымгольц пишет: «Немногочисленность и фрагментарность ископаемых находок Sepioidea затрудняет их классификацию. Границы родов устанавливаются по морфологическим особенностям раковины, систематическое значение которых не всегда ясно… Формы, характеризующие разные этапы эволюции, встречаются в одновременных отложениях» (с. 162). Напомню, Вы ссылаетесь на каталоги ХІХ в., соответствующий же том вышел в 1958 г.
Сказанное относится и к мезозойским и палеозойским находкам спирулид. Удивляет меня и Ваше замечание о продолжительности плавания раковин спирул. В открытом океане им попросту не обо что разбиться. Это не означает, что они прочны при встрече с песком и камнями. На побережье Средиземного моря я встречал, как субфоссильные залежи двустворок Glycymeris violaceus (возрастом 100 лет и более), так и практически неповрежденные, но свежие сепионы, которые, как правило, не залеживаются дольше пары недель. Со спирулой in situ дела не имел, но раковину ее держал в руках. К сожалению, она разбилась, находясь внутри пластмассовой коробки с ватой. — Против принятой версии свидетельствует морфология раковины белемнозеин. Так их рострум, несмотря на крайнюю изменчивость, не редуцируется, тогда как смена изогнутости более консервативной камерной части — фрагмокона — налицо (рис. 2).
— Ростр как раз постепенно редуцируется от палеоцена к эоцену и олигоцену.
Здесь Вы принимаете маневр в обход аргумента. Речь идет не о временнОй последовательности (как указано выше, не могущей быть надежным критерием), а о степени изогнутости фрагмокона, отличающей семейства Belemnosidae, Belopteridae и Spirulirostridae. Как можно убедиться по иллюстрации и даже по самому названию “Spirulirostra”, у рода, наиболее близкого к спируле по степени изогнутости фрагмокона, ростр прекрасно развит. —В отличие от белемнитов ростр белемнозеин, формировавшийся натянутым на раковину мантийным мешком, не развивается на вогнутой брюшной части фрагмокона (рис. 3).
Сама форма фрагмокона спирулирострид предполагает брюшное зияние. Следствие достаточно ясное, но из него следует, что выпрямленные белемнозиды происходят от искривленных, вопреки существующей версии.
Руководствуясь последней, Вы предлагаете следующее объяснение:
—у спирулид постепенно происходит смещение внутренностей (т.е. веса тела) под поплавок (фрагмокон), а противовеса, соответственно - наверх поплавка. Нижняя его часть теперь окружена не кожистой секретирующей, а нормальной мускульной мантией. Поэтому в рострах снизу образуется зияние.
Объяснение это гораздо сложнее представленного мной.
Вот, что могу возразить на него:
1. У спирулиростры с изогнутым фрагмоконом рострум прекрасно развит в нижней своей части (капитулюм), представляющей собой внешнюю сторону фрагмокона, тогда как вовнутрь он не заходит.
2. У всех колеоидей, в т. ч., у сепий и спирулы, раковина находится внутри мантийного мешка, образованного всецело секретирующей тканью.
Согласно написанному Вами, мантийные мускулы отделяют раковину от нижележащих внутренностей. На самом деле мантийная мускулатура окружает внутренностный мешок, а не отделяет его от раковины.
— Если бы указанная Вами особенность имела место, то спирулиды не могли бы строить нижнюю часть не только рострума, но и фрагмокона. Да и к чему была бы такая адаптация? Мускулы сжимали раковину снизу? Отталкивали от нее внутренности?! — Впрочем, брюшная борозда на роструме белемнитов может свидетельствовать об их происхождении от форм с зиянием, вызванным изгибом фрагмокона.
— А как быть с белемнитами, у которых борозда на спинной стороне? А если борозды есть на спинной и брюшной сторонах одновременно?
Принимаю данное возражение как возможное. Причина образования борозд нуждается в изучении. Все же, представляется вероятным, что боковые борозды служили для прикрепления мускулатуры к мантийному мешку. Брюшная же борозда, наличествующая у большинства, если не у всех белемнитов, могла возникнуть независимо от боковых и спинных, и быть унаследованной от спиральных предков. Впрочем, в данном предположении я уверен менее, чем в филогенезе спирулы. Выводу о происхождении спирулы от аммонитов Вы противопоставили следующие вопросы:
1. Где у спирулы аммонитовая линия? У прямых (гетероморфных) аммонитов - бакулитов и др. - аммонитовая линия выражена отлично!
2. Если считать, что эндогастрическая раковина спирулы повернулась в теле на 180 град. в поперечном направлении, то почему вдруг брюшной сифон аммонитов, максимально приближенный к внешнему краю, оказывается у экзогастрической спирулы на внутренней стороне спирали???
Начну со второго. Вместо того, чтобы взвесить аргумент (т. е. предположить оптимальный ответ на него), Вы приняли его за действенное возражение, лишь потому, что заметили это. На самом же деле, он свидетельствует, скорее, в пользу предложенного решения.
Ответ Вы, незаметно для себя, дали сами. Сифон реориентирован по отношению к реориентированной же раковине, но не к туловищу. Как и у большинства аммонитов, он приближен к брюшной стороне тела. N. B. задавая вопрос, Вы забыли о «меньшинстве», т. е. палеозойских климениях — наружнораковинных аммонитах со спинным сифоном. Т. о., реориентация сифона вообще не должна рассматриваться как свидетельство против аммонитного происхождения спирулы. В первом вопросе Вы рассматриваете аммонитную линию, как раз и навсегда данный признак, вне зависимости от адаптивного значения. Так, пожалуй, можно заявить, что слепыш не происходит от зрячих грызунов.
Линия образовывалась выростами мантии и служила прикреплению тела к наружной раковине. У спирулы, во-первых, раковина внутренняя; во-вторых, по сравнению с наружнораковинными аммонитами поверхность перегородок сильно уменьшена.
Это значит, что для сохранения аммонитной линии мантийные выросты должны были
а) уменьшиться в десятки раз;
б) противостоять возросшему давлению, для чего не было никакой адаптивной потребности. Напротив, редукция выростов позволяет распределить давление жидкости наиболее равномерно. Это объясняет отсутствие лопастей даже в рудиментарном виде.
Вашему вопросу о линии можно противопоставить таковой о внутреннераковинных придатках. Свой ответ Вы резюмируете словами: «Кстати, у ископаемых ростровых спирулид проостракумы найдены». Все равно, как если я бы заявил: «Кстати, у ископаемых наружнораковинных аммонитов сложная линия найдена». Вопрос поставлен не о ростровых спирулидах, а о происхождении от них спирулы; о редукции не только проостракума, но и рострума. Объяснить вторичное отсутствие таких придатков даже в рудиментарном виде (как у сепиона) сложновато. Проще предположить его первичным. В этом случае спирула должна происходить от аммонитов. Отсутствие проостракума (в отличие от перегородочной линии) Вы объясняете адаптацией: «функция проостракума - создавать твердо-эластичную продольную опору тела, что важно в первую очередь для активных пловцов. Спирула таковым не является, поэтому у нее проостракум редуцирован».
Пассивный образ жизни мог бы объяснить значительную редукцию (но не полное исчезновение) не только проостракума, но и рострума. Тем не менее, как я уже писал, спирула является активным пловцом. Об этом свидетельствует уже ее мускулистая мантия (Несис, Океанические головоногие, М. 1985, c. 191). У пассивных же организмов мускулатура редуцируется, как у кальмаров-батискафов кранхиид; желатинистых глубоководных осьминогов и мн. др.
Вы также пишете: «способность спирулы к реактивному движению на коротких промежутках времени никак не противоречит пассивному образу жизни. А ведь большую часть времени спирула именно пассивна». Из приведенного отрывка (отчета экспедиции Брууна) следует противоположное: вначале, будучи шокированными, спирулы зависли вниз головой. Потом, придя в себя, стали активно плавать в течение продолжительного времени: «Как и другие каракатицы, спирула часто производит лёгкие пульсирующие движения, внезапно бросаясь в любом направлении: вверх, вниз или из стороны в сторону... обычно животное двигалось задним концом вперёд,... прижав к нему плавники. Реже спирула... делала бросок головой вперёд» (см. приложенную фотокопию текста). Из пассивного образа жизни следует подчинение животного силам плавучести раковины, невозможность плавать в положении, отличном от вертикального. Между тем, из описания явствует, что спирула плавает не хуже других каракатиц (чего нельзя сказать о кранхиидах сравнительно с пр. кальмарами); что она с легкостью принимает горизонтальное положение и даже движется задним концом вниз, преодолевая силу плавучести раковины, а также сопротивление присосковидного светящегося органа.
Полагаю, что версия о пассивном образе жизни спирулы возникла из предрассудка, которым подменили теорию, проигнорировав достаточно ясный текст. 3 следующих недоразумения отнесу к невнимательному прочтению текста: І. —… у спирулид и аммонитов протоконх открыт, тогда как у белемноморф он отделён от прочих камер вторичной мембраной… и т. д.
— протоконх "открыт" и у ростровых третичных спирулид
В данном случае, Ваше дополнение излишне, поскольку о том же говорится и в цитате. ІІ. Вы цитируете фразу: «Такая парадоксальная тенденция как спирализация внутренней раковины, утратившей защитную функцию…», и спрашиваете: «Главная функция внутренней раковины - опорная, плюс это часть гидростатического аппарата. В чем защитная функция?»
По-видимому, торопясь ответить, Вы упустили из виду подчеркнутое слово. Факт, что внутренняя раковина происходит от наружной, первичная функция которой была защитной. ІІІ. — спиной край лучше развит у всех прямо- внутреннераковинных головоногих, в т. ч. у современных каракатиц и кальмаров. Тем не менее, раковина белемнитов также реориентирована. У их наружнораковинных предков — бактритоидей брюшной край был развит сильнее, что, в общем, соответствует первичной ориентации раковины у моллюсков, обнаруженной мною. Такая ориентация определяется положением жабр, абсорбирующих соли, идущие на постройку раковины и выделяемые через анус вследствие несовершенной выделительной системы (рис. 5). Поскольку у внутреннераковинных головоногих устье сильно уменьшено, брюшной край приближен к спинному, а жабры отдалены от обоих, метаболизм не препятствовал реориентации раковины, которая в любом случае имела место, и которую надлежит объяснить.
— вообще ничего не понял.
Я не употребил ни одного чуждого Вам термина. Если же данное суждение неверно либо усложнено, то его для начала следует постараться понять. Тем более, это следует сделать, если сложность высказывания соответствует проблеме. Вы просто ленитесь думать, а в результате науку бьют попы да гадалки. N. B. накануне Нового Года прикрепил иллюстрации. Рис. 5, полагаю, поможет Вам разобраться. О первичной ориентации я докладывал на малакологической конференции в Житомире; в Институте Зоологии в Киеве. Судя по всему, большинство слушателей меня прекрасно понимало. Похоже, что первое испытание на прочность статья прошла успешно. Надеюсь, что следующие доводы будут более обоснованными. И еще: предположите, что написанное верно. В этом случае настоящая работа представляет собой важный вклад в теорию. Можете ли Вы помочь в ее продвижении и отстаивании?
P. S. прошу прощения за возможные резкости. Полагаю, что для взвешенного ответа мне понадобилось бы пару недель (а не дней) работы.
Жду ответа,
Э. О. Хейфец
Сообщение отредактировал heyfetzed - Суббота, 09.01.2010, 00:14 |
| |
| |
| AlexIpp | Дата: Суббота, 09.01.2010, 00:10 | Сообщение # 9 |
|
Группа: Администраторы
Сообщений: 579
Репутация: 0
Статус: Offline
| Эдуард, не сочтите за нескромный вопрос:
Quote (heyfetzed) Сказанное относится и к мезозойским и палеозойским находкам спирулид Quote (heyfetzed) Факты же, добытые предшественниками, отрицать не намерен.
Отлично. А что вы знаете о мезозойских и (особенно) палеозойских находках спирулид? Хотя бы - что это за находки и на каком основании их отнесли к спирулидам?
всё о серпулидах & белемнитах
|
| |
| |
| AlexIpp | Дата: Суббота, 09.01.2010, 00:17 | Сообщение # 10 |
|
Группа: Администраторы
Сообщений: 579
Репутация: 0
Статус: Offline
| Quote (heyfetzed) палеозойских климениях — наружнораковинных аммонитах со спинным сифоном. Т. о., реориентация сифона вообще не должна рассматриваться как свидетельство против аммонитного происхождения спирулы.
да поправит меня знаток аммонитов mhorn, но мне всегда казалась логичной версия независимого происхождения климений от бактритов, за счет эндогастрического закручивания раковины.... и тогда к другим аммонитам они имеют слабое отношение...
всё о серпулидах & белемнитах
|
| |
| |
| heyfetzed | Дата: Суббота, 09.01.2010, 00:37 | Сообщение # 11 |
|
Группа: Проверенные
Сообщений: 21
Репутация: 0
Статус: Offline
| Цитата несколько вырвана из контекста. Вы противопоставили спируле мезо- и палеозойские находки спирулид. Я привел свидетельство Крымгольца из "Основ палеонтологии" о редкости сепиид вообще и "спирулид" - в частности. Об их признаках (арагонитовый рострум и открытый протоконх) я уже писал. Полагаю, что к этому следовало бы прибавить и изгиб фрагмокона - этих явно первичных признаков недостаточно для сближения Grenlandiabellitesa со "спирулидами" в противовес белемнитам. Кроме того, если спирула происходит от аммонитов, то ее следует выделить в отдельный отряд с единственным представителем, а формы с внутреннераковинными придатками отнести к отряду Belemnoseida. О поcледних, честно говоря, знаю крайне мало - изучал, в первую очередь, спирулу. Полагаю, что знаний достаточно, дабы утверждать первичность отсутствия внутреннераковинных придатков у спирулы и ее происхождение непосредственно от аммонитов. Невежеством в данной области хвастать не собираюсь, но сужу о том, что изучил досконально. Предлагаю сосредоточиться на основных положениях работы.
С уважением, Э. О. Хейфец.
|
| |
| |
| heyfetzed | Дата: Суббота, 09.01.2010, 00:45 | Сообщение # 12 |
|
Группа: Проверенные
Сообщений: 21
Репутация: 0
Статус: Offline
| По поводу климений спорить не буду. В любом случае данный случай достоин объяснения. Удовлетворил ли Вас ответ о причине реориентации сифона у спирулы?
N. b. в свое время я посвятил этому, а в особенности реориентации функциональных участков мантии целый абзац, но затем увидел, что он лишь загромождает текст (рисунок прикрепляю ниже).
Сообщение отредактировал heyfetzed - Суббота, 09.01.2010, 00:49 |
| |
| |
| heyfetzed | Дата: Суббота, 09.01.2010, 00:51 | Сообщение # 13 |
|
Группа: Проверенные
Сообщений: 21
Репутация: 0
Статус: Offline
| 
Сообщение отредактировал heyfetzed - Суббота, 09.01.2010, 01:08 |
| |
| |
| AlexIpp | Дата: Суббота, 09.01.2010, 14:17 | Сообщение # 14 |
|
Группа: Администраторы
Сообщений: 579
Репутация: 0
Статус: Offline
| давайте попробуем проще.
Есть такое понятие, как синапоморфия. Уникальная черта строения, объединяющая какую-либо эволюционную ветвь организмов. Именно на синапоморфиях обычно строятся доказательства принадлежности той или иной группы к единой эволюционной ветви. Плюс - на стратиграфических данных, которые вы отрицаете в целом
Например, синапоморфия аммонитов - сложная перегородочная линия. Спиральная раковина - нет, она независимо появляется, у наутилоидей и колеоидей как отражение тенденции к компактизации раковины (спирула:))
а какие конкретно синапоморфии у аммонитов+спирулы??? Честно говоря, не вижу никаких доказательств сходства, кроме спиральной раковины.
всё о серпулидах & белемнитах
|
| |
| |
| AlexIpp | Дата: Суббота, 09.01.2010, 14:28 | Сообщение # 15 |
|
Группа: Администраторы
Сообщений: 579
Репутация: 0
Статус: Offline
| Quote (heyfetzed) Вы противопоставили спируле мезо- и палеозойские находки спирулид
я лишь обратил внимание, что вы строите филогению без какого-либо учета данных по стратиграфическому распространению этой группы головоногих. Плюс - совершенно не знаете фактического материала (т.е. конкретных находок из мезозоя и палеозоя), описанного учёными, не желающими замечать такую очевидную и гениальную вещь, как происхождение спирулы от аммонитов.
Quote (heyfetzed) Можете ли Вы помочь в ее продвижении и отстаивании?
нет, конечно. Потому что изложенное в ней мне кажется необоснованным.
всё о серпулидах & белемнитах
|
| |
| |
| mhorn | Дата: Суббота, 09.01.2010, 20:47 | Сообщение # 16 |

Admin
Группа: Администраторы
Сообщений: 3615
Репутация: 0
Статус: Offline
| Quote (heyfetzed) выпрямлённые формы происходят от более изогнутых, а не наоборот. Такая тенденция обычна у внутреннераковинных моллюсков.
Такой тенденции у внутреннераковиных моллюсков нет, они все - потомки бактритоидей, у которых раковины была прямая
Головоногие с изогнутыми (и закручеными) раковинами происходили от прямораковинных форм неоднократно. У нижнемеловых аммонитов известны как многочисленные примеры происхождения гетероморфных аммонитов от нормально закрученных, так и происхождение мономорфных аммонитов от гетероморфных предков - три самых разнообразных и многочисленных аптских мономорфных надсемейств имеют гетероморфных предков (см. статью Михайловой и Барабошкина, 2009 - http://rogov.zwz.ru/Mikhailova,Baraboshkin,2009_Heteromorphs_rus.pdf ) Quote (heyfetzed) В случае опасности спирула втягивает голову с конечностями в мантию, которая закрывается мантийными клапанами
втягивать голову в мантию могут многие головоногие Quote (heyfetzed) Спирула оказывается не потомком белемнозенин, а представителем предковой группы, происходящей от наружнораковинных спиральных форм. К таковым относятся аммониты, для которых, как и для других потомков бактритоидей характерна зародышевая камера (протоконх), отличающая их от прочих спиральных головоногих
У всех аммонитов - и гетероморфных, и мономорфных - первый оборот закрученный. Только у некоторых раннедевонских форм это не так Quote (heyfetzed) выпрямлённые формы происходят от более изогнутых, а не наоборот. Такая тенденция обычна у внутреннераковинных моллюсков.
Такой тенденции у внутреннераковиных моллюсков нет, они все - потомки бактритоидей, у которых раковины была прямая
Головоногие с изогнутыми (и закручеными) раковинами происходили от прямораковинных форм неоднократно. Причем в палеозое имелись как эндо- так и эндогастрические наружнораковинные головоногости - произошедшие и те и другие от прямораковинных предков. У нижнемеловых аммонитов известны как многочисленные примеры происхождения гетероморфных аммонитов от нормально закрученных, так и происхождение мономорфных аммонитов от гетероморфных предков - три самых разнообразных и многочисленных аптских мономорфных надсемейств имеют гетероморфных предков (см. статью Михайловой и Барабошкина, 2009 - http://rogov.zwz.ru/Mikhailova,Baraboshkin,2009_Heteromorphs_rus.pdf ) Quote (AlexIpp) да поправит меня знаток аммонитов mhorn, но мне всегда казалась логичной версия независимого происхождения климений от бактритов, за счет эндогастрического закручивания раковины.... и тогда к другим аммонитам они имеют слабое отношение...
Поправлю. Климении - потомки гониатитов. И раковина у них, как и у всех остальных аммонитов, экзогастрическая, но сифон не совсем на месте. Впрочем, в палеозое с этим делом у аммонитов бывало по-всякому - есть и формы с почти центральным положением сифона Quote (AlexIpp) а какие конкретно синапоморфии у аммонитов+спирулы??? Честно говоря, не вижу никаких доказательств сходства, кроме спиральной раковины.
согласен, нет у них никаких синапоморфий, а закрученная раковина скорее может служить доводом против происхождения спирулы от аммонитов к разговору о хрупких не сохраняющихся раковинах спирул - прикрепляю фотографию плейстоценовой спирулы из коллекции Британского музея Естественной истории 
верхнеюрские и нижнемеловые аммониты и всё что с ними связано
|
| |
| |
| AlexIpp | Дата: Воскресенье, 10.01.2010, 02:29 | Сообщение # 17 |
|
Группа: Администраторы
Сообщений: 579
Репутация: 0
Статус: Offline
| Ну и пройдемся по мелочам.
Хотя, мне было бы интересней получить четкие ответы на вопросы по сути (про синапоморфии и как увязывается ваша идея со стратиграфическим распространением ископаемых - т.е. с фактическим материалом). Но я прекрасно понимаю, что пытаться разубедить человека в идеях, которые он вынашивал длительное время, практически бесполезно. Quote (heyfetzed) Напомню, Вы ссылаетесь на каталоги ХІХ в., соответствующий же том вышел в 1958 г
я ссылаюсь на каталоги 19 века, так как в них спирулиды упоминаются регулярно - а это и есть показатель того, что они совсем не редкие. По-настоящему редкие формы в каталоги 19 в попадали нечасто  Quote (heyfetzed) Это не означает, что они прочны при встрече с песком и камнями
и тем не менее, ископаемые находки спирул - есть и они отделены от последних аммонитов 50 миллионами лет. Quote (heyfetzed) У всех колеоидей, в т. ч., у сепий и спирулы, раковина находится внутри мантийного мешка, образованного всецело секретирующей тканью. Согласно написанному Вами, мантийные мускулы отделяют раковину от нижележащих внутренностей. На самом деле мантийная мускулатура окружает внутренностный мешок, а не отделяет его от раковины мантия и мускулистая мантия - это разные вещи. Разбирайтесь. Мускулистая мантия окружает полукольцом все мягкое тело и сверху крепится к краям проостракума (пока он был). У ростровых спирулид - частично и к боковым краям ростра.
Про наличие мускулов снизу между раковиной и мягким телом я не писал, откуда вы это взяли?
Quote (heyfetzed) Да и к чему была бы такая адаптация? Мускулы сжимали раковину снизу? Отталкивали от нее внутренности?!
сжимали бы мантийную полость, выталкивая из нее воду через воронку. Такова ее функция у всех колеоидей.
Quote (heyfetzed) Сифон реориентирован по отношению к реориентированной же раковине, но не к туловищу
а почему бы не наоборот? Расположение органов мягкого тела у всех моллюсков - более пластичный признак, чем расположение элементов раковины.
Да. А почему в онтогенезе или строении спирулы нет следов поворота раковины? Подобно тому, как есть следы поворота раковины у гастропод? Про аммонитовую линию:
Quote (heyfetzed) противостоять возросшему давлению, для чего не было никакой адаптивной потребности. Напротив, редукция выростов позволяет распределить давление жидкости наиболее равномерно
о каком давлении идет речь? тела любых глубоководных животных не содержат воздушных полостей тела, гидростатическое давление им по фигу - их тела практически несжимаемы. Другой вопрос, что у спирулы есть фрагмокон, заполненный газом.... и для упрочнения этого фрагмокона наличие аммонитовой линии было бы очень кстати
Quote (heyfetzed) Пассивный образ жизни мог бы объяснить значительную редукцию (но не полное исчезновение) не только проостракума, но и рострума.
совершенно не обоснованное утверждение Quote (heyfetzed) Я не употребил ни одного чуждого Вам термина.
попробую объяснить:
Quote (heyfetzed) спиной край лучше развит у всех прямо- внутреннераковинных головоногих
что конкретно имеется в виду под словами "лучше развит"? Quote (heyfetzed) Такая ориентация определяется положением жабр, абсорбирующих соли, идущие на постройку раковины и выделяемые через анус вследствие несовершенной выделительной системы
поясните. В чем функциональная связь ануса с раковиной??? Quote (heyfetzed) Поскольку у внутреннераковинных головоногих устье сильно уменьшено
а где у них устье? И что значит - уменьшено? А как бы оно выглядело, если бы было "не уменьшено?
Quote (heyfetzed) брюшной край приближен к спинному
это как? они сильно сплющены в спинно-брюшном направлении?Quote (heyfetzed) а жабры отдалены от обоих, метаболизм не препятствовал реориентации раковины
вот это интересно. А как метаболизм ограничивает положение раковины?
всё о серпулидах & белемнитах
|
| |
| |
| AlexIpp | Дата: Воскресенье, 10.01.2010, 02:33 | Сообщение # 18 |
|
Группа: Администраторы
Сообщений: 579
Репутация: 0
Статус: Offline
| Quote (mhorn) Климении - потомки гониатитов
этот переход прослежен на каменном материале?
всё о серпулидах & белемнитах
|
| |
| |
| mhorn | Дата: Воскресенье, 10.01.2010, 16:53 | Сообщение # 19 |
|
Admin
Группа: Администраторы
Сообщений: 3615
Репутация: 0
Статус: Offline
| Quote (AlexIpp) этот переход прослежен на каменном материале?
насколько я понимаю - да
вот что пишет Ежи Дзик (Dzik, 2006, Paleont.Pol., no.63, c.311): "There is little doubt that clymenias originated from Protornoceras (House 1970) and that the South Urals Kirsoceras is the connecting link. Yet, the exact course of the transition at the ordinal boundaries has not been determined. The protornoceratids form a whole plexus of species differing in the evoluteness of the conch and usually showing remarkable population variability. They continue their occurrence throughout the C. quadrantinidosa, C. marginifera, and the early P. trachytera zones, probably represented by lineages evolving (and speciating?) at place. Their diversity gradually increased and the immigration of the first clymenias at the beginning of the P. trachytera Zone is hardly discernible in the spectrum of the ammonoid conch forms. This is only the location of the siphuncle (and replacing the ventral lobe with dorsal one) which makes the difference."
верхнеюрские и нижнемеловые аммониты и всё что с ними связано
|
| |
| |
| heyfetzed | Дата: Понедельник, 11.01.2010, 00:48 | Сообщение # 20 |
|
Группа: Проверенные
Сообщений: 21
Репутация: 0
Статус: Offline
| Уважаемые оппоненты!
Время не позволит мне ответить на все заданные вопросы, но по возможности, постараюсь учесть максимум.
AllIpp, Вы пишете, якобы я не признаю стратиграфические данные. На самом деле, Вы не желаете признавать недостаток стратиграфических данных для эмпирической реконструкции. В данном случае о них свидетельствует беспристрастное и авторитетное издание. Цитату я привел.
Сравнительно редкие ископаемые дают лишь "отрицательный" ответ: данная группа не может появиться позже либо вымереть раньше времени жизни находки. Филогенез в таком случае следует восстанавливать теоретически. N. b. замечание о т. н. "гениальности" эмпирично: не могли же поколения ученых заблуждаться и оставлять недоработки в теории эволюции! История свидетельствует о противном. Примеров достаточно.
N. b. Таково может быть истинное основание Вашего оппонирования. Если бы спирулу изначально произвели от аммонитов (пусть даже ошибочно), Вы бы приняли это за истину и так же спорили с тем, кто производит ее от белемнитоподобных форм.
Простите, но судя по ответу на цитату: "Пассивный образ жизни мог бы объяснить значительную редукцию (но не полное исчезновение) не только проостракума, но и рострума". - "совершенно не обоснованное утверждение", - Вы считаете себя, если не гением, то совершенным критерием обоснованности. Не менее "обоснованным" (вернее авторитарным) представляется и высказывание mhorn'a: "а закрученная раковина скорее может служить доводом против происхождения спирулы от аммонитов". Конец цитаты(!) О недостаточной теоретической подготовке свидетельствуют Ваши вопросы о положении сифона и лопастной линии, а также ответ по поводу отсутствия внутреннераковинных придатков у спирулы - он остается в силе: Происхождению спирулы от белемнитоподобных форм, противоречит отсутствие даже в рудиментарном виде рострума, крыльев и проостракума.
Вам следует повысить уровень теоретизации. N. b. в связи с этим буду рад ответить на вопросы по первичной ориентации раковины у моллюсков.
- Выпрямлённые формы происходят от более изогнутых, а не наоборот. Такая тенденция обычна у внутреннераковинных моллюсков.
- Такой тенденции у внутреннераковиных моллюсков нет, они все - потомки бактритоидей, у которых раковины была прямая.
Головоногие с изогнутыми (и закручеными) раковинами происходили от прямораковинных форм неоднократно.
-Внутреннераковинными моллюсками являются не только головоногие, но и брюхоногие, например, полуслизень Daudebaria, слизни с седловидным рудиментом раковины, переднежаберный моллюск Sigaretus (название говорит само за себя), родственный Natica, крепкая раковина которой напоминает виноградную улитку и т. д. Следующий тезис: "Они все потомки бактритоидей у которых раковина была прямая" оспаривается; следовательно, сам нуждается в обосновании. Повторюсь: отсутствие внутреннераковинных придатков у спирулы свидетельствует против ее происхождения от белемнитоподобных форм.
Наконец, Вы "опровергаете" меня примером, с которым не могу поспорить. Ведь у данных головоногих раковина была наружной. В цитате же речь идет о раковинах внутренних! Повторюсь, Вы не взвешиваете доводы. По поводу замечания об аммонитной линии. Вы пишете о синапоморфии аммонитов. Допустите, что спирула происходит от аммонитов, и утратила сложнорассеченную линию вторично. Значит ли это, что ее нельзя относить к аммонитам? Значит ли, что кита или змею нельзя относить к четвероногим, как это принято в зоологии позвоночных?
Насколько мне известно, функцией мантийной линии считается фиксация тела к наружной раковине. У раковины спирулы эта функция отстутвует. Значит, основания для редукции налицо.
-Другой вопрос, что у спирулы есть фрагмокон, заполненный газом.... и для упрочнения этого фрагмокона наличие аммонитовой линии было бы очень кстати.
-Строящиеся перегородки заполнены жидкостью, давление которой и предопределяет форму прилегающих участков мантии. Что до упрочнения раковины, то поскольку она окружена по-преимуществу мягкой тканью; поскольку спирула не встречается в прибойной зоне; наконец, поскольку раковина не защищает спирулу от хищников, такое упрочнение было бы излишеством. Перехожу к Вашим вопросам о первичной ориентации раковины:
-спиной край лучше развит у всех прямо- внутреннераковинных головоногих
-что конкретно имеется в виду под словами "лучше развит"?"
Может, я не совсем удачно выразился. Спиной край раковины длинее брюшного. -Такая ориентация определяется положением жабр, абсорбирующих соли, идущие на постройку раковины и выделяемые через анус вследствие несовершенной выделительной системы
-поясните. В чем функциональная связь ануса с раковиной?"
-Пояснение содержится в тексте. Тем не менее готов расписать это несколько подробнее: Занимаясь причиной торсиона у брюхоногих, я обратил внимание на то, что, как показано на рис. 5, задний край раковины моллюсков, как правило, длинее переднего.
Сзади у моллюсков находится анус, выделяющий ядовитые отходы организма и жабры, абсорбирующие жизненно необходимые вещества. Такое странное соседство можно объяснить тем, что анус выделяет избыток солей, а жабры их абсорбируют. Соли идут на постройку раковины. В результате задний ее край растет быстрее переднего. В руководстве по физиологии животных (Wilson J. A. Principles of animal physiology [p. 603]. The Macmillan company, New York, 1972. – 891 р.) я нашел подтверждение такому выводу. -Поскольку у внутреннераковинных головоногих устье сильно уменьшено
-а где у них устье? И что значит - уменьшено? А как бы оно выглядело, если бы было "не уменьшено?
-Устье наличествует у колеоидей с замкнутым фрагмоконом. О дальнейшем, см. ответ к следующему вопросу. -брюшной край приближен к спинному
-это как? они сильно сплющены в спинно-брюшном направлении?
-Нет, уменьшен диаметр фрагмокона по отношению к таковому наружнораковинных головоногих. Тем не менее, поскольку вопрос заключается в соотношении роста спинного и брюшного краев раковины, в данном случае существенно их сближение между собой. -а жабры отдалены от обоих, метаболизм не препятствовал реориентации раковины
-Вот это интересно. А как метаболизм ограничивает положение раковины?
-Задавая последний вопрос, Вы успели позабыть второй (на эту тему). См. ответ на него и рис. 5. к статье. Закончу цитатой, имеющей отношение к торсиону у брюхоногих: -Сифон реориентирован по отношению к реориентированной же раковине, но не к туловищу
-а почему бы не наоборот? Расположение органов мягкого тела у всех моллюсков - более пластичный признак, чем расположение элементов раковины.
Да. А почему в онтогенезе или строении спирулы нет следов поворота раковины? Подобно тому, как есть следы поворота раковины у гастропод?
-Потому, что данный случай уникален: не эволюционная перестройка раковины, но внезапный ее поворот, ведущий к подъему центра плавучести (напомню, что данный довод оказался неразобранным).
По поводу следов поворота:
1. В отличие от переднежаберных или перекрестнонервных (Streptyneura), у заднежаберных и легочных брюхоногих, составляющих группу Euthyneura следы поворота раковины исчезают (хотя у первых сохраняются в ходе личиночного развития). То же могло иметь место и у спирулы.
2. Уверены ли Вы, что и в онтогенезе, и в анатомии спирулы на всех ее стадиях не сохранилось никакого следа данного явления, тем более, что о нем никто до сих пор не догадывался, следовательно, и не искал? Препарировали ли Вы спирулу? Похоже, что в данном случае Вы выдаете отсутствие достоверных знаний о признаке за достоверные знания об его отсутствии.
До связи,
Эдуард. P. s. AllIpp пишет, что втягивать голову в раковину могут многие головоногие. Один из рецензентов уже приводил такое замечание. У Несиса я обнаружил ссылку на один вид эгопсидных кальмаров, втягивающий голову лишь частично и в молодом возрасте, кроме того каракатица способна втягивать основание головы под языковидный выступ сепиона (но не вовнутрь мантии).
Ссылкой в данный момент не располагаю. См. Несис, "Краткий определитель головоногих", М. 1982.
Сообщение отредактировал heyfetzed - Понедельник, 11.01.2010, 01:00 |
| |
| |
| AlexIpp | Дата: Понедельник, 11.01.2010, 01:01 | Сообщение # 21 |
|
Группа: Администраторы
Сообщений: 579
Репутация: 0
Статус: Offline
| Попробуем "укрупнить" предмет нашей дискуссии и не влезать в обсуждение деталей - в данном случае они имеют малое значение и уводят от главного. Наука - это не попытка найти доказательства какой-то идеи и любой ценой вывернуться из противоречий.
Наука - это поиск наиболее непротиворечивого объяснения существующим фактам. Либо проверка идей путем фальсификации (не подумайте плохого, это термин из философии Поппера) - "если бы это было правильно, то из этого бы следовало..." А если не следует, то идея отбрасывается. Поэтому наука начинается со сбора всех фактов, продолжается - научным анализом и фальсификациями гипотез, а заканчивается - выводами. По-моему, изложенное выше содержит явные проблемы с первой и второй процедурами. Вы стараетесь найти доказательства идеи, в которую верите, не замечая, что реальных аргументов (тех самых синапоморфий) у вас нет.
всё о серпулидах & белемнитах
|
| |
| |
| mhorn | Дата: Понедельник, 11.01.2010, 01:11 | Сообщение # 22 |
|
Admin
Группа: Администраторы
Сообщений: 3615
Репутация: 0
Статус: Offline
| Quote (heyfetzed) Сравнительно редкие ископаемые дают лишь "отрицательный" ответ: данная группа не может появиться позже либо вымереть раньше времени жизни находки
Ещё раз: это далеко не самые редкие ископаемые. Ознакомьтесь, пожалуйста, с другой литературой по теме, кроме "Основ палеонтологии". То, что Н.Г.Крымгольц писал о редкости спирулид, на мой взгляд, связано с тремя причинами: 1) во-первых, "Основы палеонтологии" основное внимание уделяли тем таксонам, находки которых были к этому времени известны в СССР 2) во-вторых, Крымгольц был специалистом по мезозойским головоногим и никогда не изучал кайнозойских представителей группы 3) в третьих, как следствие из п.2, по сравнению с обилием и разнообразием мезозойских головоногих в кайнозое они, конечно, редки Quote (heyfetzed) Происхождению спирулы от белемнитоподобных форм, противоречит отсутствие даже в рудиментарном виде рострума, крыльев и проостракума.
ну почему же? то, что у аргонавтов не сохранилось и следа раковины, не мешает относить их к осьминогам По поводу происхождения спирулы (а заодно эволюции других колеоидей) Вам, полагаю, будет интересно прочитать автореферат В.А. Бизикова, ведущего специалиста по современным головоногим (он опубликовал также большую. книгу на эту тему, но в электронном виде её у нас нет). вот ссылка: http://rogov.zwz.ru/Bizikov2008_autoreferat.pdf
верхнеюрские и нижнемеловые аммониты и всё что с ними связано
|
| |
| |
| AlexIpp | Дата: Понедельник, 11.01.2010, 01:25 | Сообщение # 23 |
|
Группа: Администраторы
Сообщений: 579
Репутация: 0
Статус: Offline
| Quote (heyfetzed) На самом деле, Вы не желаете признавать недостаток стратиграфических данных для эмпирической реконструкции. В данном случае о них свидетельствует беспристрастное и авторитетное издание. Цитату я привел. Сравнительно редкие ископаемые дают лишь "отрицательный" ответ: данная группа не может появиться позже либо вымереть раньше времени жизни находки. Филогенез в таком случае следует восстанавливать теоретически именно потому, что я хорошо представляю объем фактического материала по ископаемым спирулидам и его стратиграфию, я утверждаю: существующая реконструкция филогении спирулид для кайнозоя является эмпирической.
Quote (heyfetzed) Повторюсь: отсутствие внутреннераковинных придатков у спирулы свидетельствует против ее происхождения от белемнитоподобных форм.
кстати, хорошо известно, что у спирулы задний конец раковины покрыт тонким известковым слоем, который, как считается, представляет из себя РУДИМЕНТ ростра.
Quote (heyfetzed) Насколько мне известно, функцией мантийной линии считается фиксация тела к наружной раковине. У раковины спирулы эта функция отстутвует. Значит, основания для редукции налицо.
я не аммонитчик (Миша, ау?), но основной версией функции аммонитовой линии, насколько мне известно, считается именно упрочнение фрагмокона, заполненного газом.
Кстати, есть такие прямые верхнемеловые аммониты с внутренней раковиной - полиптихиты - вот у них аммонитовая линия вполне нормальная
всё о серпулидах & белемнитах
|
| |
| |
| heyfetzed | Дата: Понедельник, 11.01.2010, 01:46 | Сообщение # 24 |
|
Группа: Проверенные
Сообщений: 21
Репутация: 0
Статус: Offline
| По поводу укрепления стенки перегородочной линией я дал ответ.
Мнение Крымгольца высказано в авторитетном издании и, надо полагать, прошло проверку редколлегии.
Что касается укрупнения дискуссии, то, похоже, это первый довод, с которым готов согласиться.
В ближайшие дни постараюсь резюмировать основные вопросы. Предлагаю сделать небольшой перерыв, чтобы не увязнуть в мелочах.
Спокойной ночи!
Эдуард.
|
| |
| |
| AlexIpp | Дата: Понедельник, 11.01.2010, 02:16 | Сообщение # 25 |
|
Группа: Администраторы
Сообщений: 579
Репутация: 0
Статус: Offline
| Quote (heyfetzed)
Мнение Крымгольца высказано в авторитетном издании и, надо полагать, прошло проверку редколлегии.
к такого рода утверждениям все же лучше относиться с осторожностью.
Например, у нас на сайте выложена книга Г.К. Кабанова "Скелет белемнитид":
http://rogov.zwz.ru/Kabanov,1967_Belemnitida.pdf
Она тоже прошла рецензию и редколлегию. И что - теперь ученые всего мира обязаны взять на вооружение точку зрения о прижизненно-эластичном ростре белемнитов?
По поводу "Основ палеонтологии" mhorn все правильно написал в сообщении #22. Этот исследователь делал этот вывод по литературным данным, сам спирулид не изучал. Плюс, после выхода "Основ палеонтологии" были опубликованы находки многих родов из палеозоя и мезозоя - вот (неполный? - по памяти) список для обсуждения: Adygeia, Groenlandibelus, Belospirula, Kostromateuthis, Shimanskya, Naefia... Плюс новые находки собственно спирул - в миоцене (что моложе всех перечисленных и моложе всех третичных ростровых спирулид). А вы говорите - "недостаточно данных". Достаточно...
всё о серпулидах & белемнитах
|
| |
| |
| mhorn | Дата: Понедельник, 11.01.2010, 10:47 | Сообщение # 26 |
|
Admin
Группа: Администраторы
Сообщений: 3615
Репутация: 0
Статус: Offline
| Quote (AlexIpp) я не аммонитчик (Миша, ау?), но основной версией функции аммонитовой линии, насколько мне известно, считается именно упрочнение фрагмокона, заполненного газом.
о функции аммонитовых сложнорассеченных перегородок до сих пор спорят. Что можно сказать точно - что у настоящих океанических аммоноидей (филло- и литоцератид) линия была сложная
Quote (AlexIpp) Кстати, есть такие прямые верхнемеловые аммониты с внутренней раковиной - полиптихиты
в смысле - птихоцерасы Quote (AlexIpp) - вот у них аммонитовая линия вполне нормальная
да, также как и у Gaudriceras, для которых также предполагается, что раковина облекалась мантией. Кстати, у аммонитов ещё очень консервативное строение первых двух перегородок - которое, само собой, очень отличается от того, что мы видим у спирулы
верхнеюрские и нижнемеловые аммониты и всё что с ними связано
|
| |
| |
| AlexIpp | Дата: Понедельник, 11.01.2010, 12:43 | Сообщение # 27 |
|
Группа: Администраторы
Сообщений: 579
Репутация: 0
Статус: Offline
| Quote (mhorn) в смысле - птихоцерасы
да, в смысле птихоцерасы...
всё о серпулидах & белемнитах
|
| |
| |
| heyfetzed | Дата: Понедельник, 18.01.2010, 13:20 | Сообщение # 28 |
|
Группа: Проверенные
Сообщений: 21
Репутация: 0
Статус: Offline
| Уважаемый Алексей Павлович,
За эти дни понял, что недооценил внутреннее обоснование существующей версии. Подробнее об этом напишу позднее. Пока же вернусь к вопросу об ископаемых остатках.
Слова Г. Я. Крымгольца резюмируют приведенную в "Основах" полную сводку родов, и отражают положение дел на тот момент (т. е. ч-з сто с лишним лет с начала изучения белемнозеин). К тому времени версия об эволюции белемнозеин вполне укоренилась в науке.
По-моему, в противовес моим словам о недостатке ископаемых для непосредственной реконструкции, у Вас возникло представление о том, что филогенез т. н. спирулид хорошо обоснован эмпирически. Достоверно Вы смогли сослаться на сравнительно поздние находки спирулы.
По поводу реплики mhorn’а и приложенной им фотографии. Не отрицаю, что хрупкие раковины спирул имеют шанс сохраниться в ископаемом состоянии — сохраняются же медузы в Венде. К тому же приведенная фотогафия битого экземпляра из сравнительно молодых плейстоценовых отложений, скорее, свидетельствует в пользу хрупкости раковины, чем наоборот.
Постараюсь сформулировать оспариваемое положение более четко: раковина спирулы не отличается от таковых белемнозеин особыми свойствами, позволяющими ей лучше сохраняться в ископаемом состоянии; в то же время у нее отсутствует ростр, который у белемноморф сохраняется наилучшим образом. Т. о., если белемнозеины редки (о чем свидетельствуют признанные вами бассейны б. СССР, изобиловавшие головоногими), тем более редкой должна быть спирула.
От белемнитов родственники спирулы были выведены уже первыми исследователями — Деге (Deshayes), Блэнвиль (Blainville), Вольц (Voltz), Орбиньи (Orbigny), — работавшими в первых десятилетиях ХІХ в. Очевидность сходства не требует строгих теоретических построений (ср. с пословицей «докажи, что ты не верблюд»). Очевидно и то, что классическая версия не подвергалась сомнению, а значит, не проходила испытания на прочность. Более того, специализированные уникальные образования представляются значительно более весомыми эволюционными признаками нежели спиральная форма раковины, возникавшая неоднократно не только у головоногих, и даже не только у моллюсков, к тому же, закрученная в обратном направлении, нежели у аммонитов либо наутилид. Подчеркнутому тексту соответствует и аргумент Алексея Пaвловича от 09.01.2010, 20:47 про неоднократное появление спирали у головоногих (не совсем кстати противопоставленный моему высказыванию о невозможности спирализации внутренней раковины). Т. о., спирализация раковины белемнозеин представляется обоснованной целым комплексом фактов. Обратное предположение требует более тщательной аргументации.
Воспользуюсь подсказкой Алексея Павловича, что спирализация раковины возникла как средство ее компактизации. С этим утверждением я согласен полностью. Другое дело, что с ним не согласуется принятая версия происхождения белемнозеин. Раковина выпрямленных белоптерид не менее компактна, чем у спирулирострид. Согласно принятой версии, здесь имеет место искривление фрагмокона наряду с увеличением его объема. Из самой функции фрагмокона напрашивается вывод о том, что спирализация увеличивает плавучесть животного. Как я уже говорил, против этого свидетельствует та же спирулиростра с ее массивным рострумом.
К тому же со всех точек зрения более эффективным средством повышения плавучести является первоначальная редукция рострума (начиная с белемнозид и белоптерид, и не ограничиваясь одной лишь спирулой). Прежде всего, рострум значительно более изменчив, чем фрагмокон, значит, такая тенденция более вероятна, чем описываемая.
Далее, редукция рострума дала бы существенную экономию материала плюс увеличение полезного объема внутри тела, и наоборот: продукция фрагмокона приведет к несущественному увеличению затрат на постройку раковины и к уменьшению полезного объема полости.
Т. о., положение о развитии фрагмокона не согласуется ни с потребностями животного, ни с относительной изменчивостью частей раковины. Поскольку группа все же эволюционировала, следует предположить обратную тенденцию, редукцию фрагмокона в результате приспособления к активному образу жизни, расставляющую все на свои места.Такому предположению соответствует брюшное зияние рострума у наименее изогнутых форм, отличающее белемнозеин от белемнитов, и связанное с изначальным изгибом фрагмокона.
Вписывается в эту тенденцию и спирула с ее отсутствием внутреннераковинных придатков, апофилией белемноморф, не менее существенной, чем лопастная линия аммонитов. Алексей Павлович замечает, что на задней части раковины спирулы имеется тонкий известковый слой, интерпретируемый, как остаток рострума. Тем не менее, таковой заметен лишь при специальном исследовании, дедуцирующем рудимент рострума из версии происхождения спирулы от белемнитоподобных организмов (на фотографии разреза раковины, прикрепленной Алексеем Павловичем 27.12.2009, 11:55 трудно обнаружить нечто существенное). За рудимент рострума мог быть принят его зачаток, предпосылка его возникновения, следующая из постоянного контакта внешней части оборота с мантийным мешком. Для сравнения, шип сепиона, распределенный по спирали, дал бы приостренный киль. Кроме того, рудиментом рострума являются крылья, характерные для сепиид и белемнозеид. Если бы они наличествовали у спирулы, фрагмокон окружала бы инволютная раковина, подобная таковой наутилуса. Не следует забывать и проостракум. Таких явных рудиментов раковина спирулы не несет, что позволяет предположить первичное их отсутствие.
Более того, как свидетельствуют наблюдения Брууна, ни рострум, ни проостракум не помешали бы спируле. Насколько мне помнится, ученый наблюдал ее в теч. 48 ч., и, согласно его свидетельствам, спирула бОльшую часть времени проводила в активном движении, преодолевая при этом силу плавучести раковины. На отчет Брууна ссылаются поздние авторы, констатируя пассивный образ жизни спирулы (см. Несис К. Н., Краткий определитель головоногих моллюсков). Интуитивно обоснованное положение о пассивном образе жизни спирулы обратилось в предрассудок, помешавший осмыслить данный пункт отчета. Подобное превратное прочтение не является уникальным. Так, гениальные опыты Спалланцани и Жюрина, доказывавшие эхолокацию у летучих мышей, авторитарно отверг отец палеонтологии Ж. Кювье, предположивший, что зверьки ориентируются с помощью дальнего осязания. В «Жизни животных» Брема под редакцией А. Н. Северцова, (1941 г.) сказано, якобы в опытах Спалланцани летучие мыши превосходно летали с закрытыми глазами и ушами.
Об активном образе жизни спирулы свидетельствуют и такие черты, как мускулистая мантия, нехарактерная для пассивных головоногих, а также светящийся орган, средство дальней коммуникации, чья функция закономерно объявлена загадочной. К последнему замечанию Алексея Павловича от 27.12.2009, 19:29: расстояние м. глазами спирулы, следовательно, диаметр окружности, проведенной через них, гораздо меньше продольной оси тела. Т. о., поле зрения спирулы недостаточно для рассмотрения светящегося органа соседней особи, устроенного, к тому же, по принципу потайного фонаря. Для обсуждения предлагаю следующие пункты: 1. Изначально белемнозеины были сближены с белемнитидами на основании достаточно глубокого сходства при сравнительно бедном ископаемом материале.
2. Такое сближение представляется внутренне обоснованным, в то время, как сходство спиральной раковины спирулы с таковой аммонитов, — поверхностно-внешним.
3. Спирализация фрагмокона, вытекающая из существующей версии, при более внимательном рассмотрении не соответствует, как потребности белемнозеин, так и относительной изменчивости частей их раковины.
4. Резонно предположить редукцию спирали у белемнозеин в пользу чего свидетельствует зияние рострума даже у наиболее выпрямленных форм.
5. В этом случае спирула должна быть предком белемнитоподобных форм, и происходить от наружнораковинных спиральных предков. Таковыми могут быть только аммониты.
6. В пользу этого свидетельствует
а) отсутствие явных рудиментов внутреннераковинных придатков у спирулы при ее активном образе жизни;
б) сходство спирулы с наружнораковинными организмами, вкл. размеры раковины, выступающие за контуры тела и пассивную защитную реакцию;
7. Обратная ориентация спирали у спирулы, равно как и обратное соотношение сторон у бактритоидей и белемноморф вызвано смещением ценра плавучести раковины книзу с последующим ее переворотом.
Жду откликов,
С уважением, Эдуард
|
| |
| |
| heyfetzed | Дата: Понедельник, 18.01.2010, 13:27 | Сообщение # 29 |
|
Группа: Проверенные
Сообщений: 21
Репутация: 0
Статус: Offline
| P. s. находясь в цейтноте, я не просматривал позднейшие сообщения. Прошу извинить, Эдуард.
|
| |
| |
| heyfetzed | Дата: Понедельник, 18.01.2010, 13:29 | Сообщение # 30 |
|
Группа: Проверенные
Сообщений: 21
Репутация: 0
Статус: Offline
| P. p. s. изображения птихоцераса пока не нашел, но с этой реконструкцией знаком, и даже один раз сослался на нее. По-моему, желаемое здесь выдают за действительное. Представьте себе, как могла проворачиваться удлиненно-элиптическая спираль внутри тела в процессе роста.
|
| |
| |
| AlexIpp | Дата: Понедельник, 18.01.2010, 19:21 | Сообщение # 31 |
|
Группа: Администраторы
Сообщений: 579
Репутация: 0
Статус: Offline
| Здравствуйте, Эдуард Quote (heyfetzed) От белемнитов родственники спирулы были выведены уже первыми исследователями — Деге (Deshayes), Блэнвиль (Blainville), Вольц (Voltz), Орбиньи (Orbigny), — работавшими в первых десятилетиях ХІХ в. Очевидность сходства не требует строгих теоретических построений (ср. с пословицей «докажи, что ты не верблюд»). Очевидно и то, что классическая версия не подвергалась сомнению
по моим представлениям, Крымгольц был одним из последних, кто выводил спирулид от белемнитов.
Был такой американский исследователь, Ю.А. Елецкий, в 1966 году он написал, наверное, самую цитируемую после Нэфа работу по ископаемым колеоидеям в XX веке. В ней он показал, что строение протоконха спирулы архаично (имеется цекум и просифон) и резко отличается от строения протоконха аулакоцератид и белемнитов. Я думаю, вам будет интересно посмотреть на эту работу. Там же детально описан гренландибелус:
Jeletzky J.A. 1966. Comparative morphology, phylogeny, and classification of fossil Coleoidea // The University of Kansas Paleontological Contributions. No. 42. Mollusca. Article 7. P. 1-162.
http://rghost.ru/853150
Большинство исследователей приняло эту аргументацию и спирулиды (и ростровые, и нормальные) с тех пор считаются независимой от белемнитов ветвью колеоидей. Альтернативную гипотезу предложили в 1999 году Хьюитт и Ягт - вторичное происхождение цекума и просифона, как результат перехода к глубоководному образу жизни (а ростровые спирулиды, по их мнению, происходят непосредственно от белемнитов).
Quote (heyfetzed) пиральная форма раковины, возникавшая неоднократно не только у головоногих, и даже не только у моллюсков, к тому же, закрученная в обратном направлении, нежели у аммонитов либо наутилид.
зато она закручена в том же направлении, что начинают закручиваться фрагмоконы ростровых спирулид
Quote (heyfetzed) Воспользуюсь подсказкой Алексея Павловича, что спирализация раковины возникла как средство ее компактизации. С этим утверждением я согласен полностью. Другое дело, что с ним не согласуется принятая версия происхождения белемнозеин. Раковина выпрямленных белоптерид не менее компактна, чем у спирулирострид. Согласно принятой версии, здесь имеет место искривление фрагмокона наряду с увеличением его объема. Из самой функции фрагмокона напрашивается вывод о том, что спирализация увеличивает плавучесть животного. Как я уже говорил, против этого свидетельствует та же спирулиростра с ее массивным рострумом.
спиральный фрагмокон дает возможность сместить центр плавучести вперед. Если при этом ростр сохраняется - это лишь означает, что функция противовеса была на этом этапе еще нужна. Заметьте, просуществовали ростровые формы со спиральным фрагмоконом очень недолго - сразу, в момент ростр исчезнуть не мог. Что в принципе логично увязывается с вашим абзацем, выделенным жирным.
Quote (heyfetzed) Поскольку группа все же эволюционировала, следует предположить обратную тенденцию, редукцию фрагмокона в результате приспособления к активному образу жизни, расставляющую все на свои места.
в общем, если описывать кратко, то ситуация понимается учеными так. После великого вымирания и исчезновения белемнитов в конце мела освободилась масса экологических ниш в мелководных сообществах, которые были заняты организмами, которые успели это сделать первыми. Редкие спирулиды, пережившие вымирание, освоили и дали вспышку разнообразия ростровых форм в палеогене. Только вот чуть позже они были довольно быстро вытеснены из этих ниш уже современными модификациями внутреннераковинных - более совершенными кальмарами, осьминогами и особенно сепиидами. Естественно, редукция ростра (даже если она была нужна) не могла произойти враз, и потому полной редукции ростра у ископаемых спирулид нет.
Quote (heyfetzed) принят его зачаток, предпосылка его возникновения, следующая из постоянного контакта внешней части оборота с мантийным мешком
исследования Бизикова, на которые уже ссылался выше mhorn, однозначно показали, что на контактах внутренней раковины и мягкого тела всегда у всех групп головоногих образуется хрящ. А не минеральные образования Quote (heyfetzed) расстояние м. глазами спирулы, следовательно, диаметр окружности, проведенной через них, гораздо меньше продольной оси тела. Т. о., поле зрения спирулы недостаточно для рассмотрения светящегося органа соседней особи, устроенного, к тому же, по принципу потайного фонаря.
предлагаю для сравнения посмотреть на картинку. Этот вопрос (радиус обзора) - классика...
http://nplit.ru/news/item/f00/s02/n0000213/pic/000003.jpg
Комментировать ничего не буду.
всё о серпулидах & белемнитах
|
| |
| |
| AlexIpp | Дата: Понедельник, 18.01.2010, 19:38 | Сообщение # 32 |
|
Группа: Администраторы
Сообщений: 579
Репутация: 0
Статус: Offline
| Quote (heyfetzed) Для обсуждения предлагаю следующие пункты:
ок.
Quote (heyfetzed) 1. Изначально белемнозеины были сближены с белемнитидами на основании достаточно глубокого сходства при сравнительно бедном ископаемом материале.
да, согласен
Quote (heyfetzed) 2. Такое сближение представляется внутренне обоснованным, в то время, как сходство спиральной раковины спирулы с таковой аммонитов, — поверхностно-внешним.
да, согласен. С дополнением, что после работы Елецкого 1966 года почти никто не выводит спирулу от белемнитов (см. выше)
Quote (heyfetzed) 3. Спирализация фрагмокона, вытекающая из существующей версии, при более внимательном рассмотрении не соответствует, как потребности белемнозеин, так и относительной изменчивости частей их раковины.
не согласен. Спирализация даже у ростровых форм несколько увеличивает расстояние между центром плавучести и задним центром тяжести. Как результат, на построение противовеса идет меньшее количество материала, что выгоднее энергетически.Quote (heyfetzed) 4. Резонно предположить редукцию спирали у белемнозеин в пользу чего свидетельствует зияние рострума даже у наиболее выпрямленных форм.
5. В этом случае спирула должна быть предком белемнитоподобных форм, и происходить от наружнораковинных спиральных предков. Таковыми могут быть только аммониты.
в этих двух положениях я ничего не понял. Спираль - это фрагмокон, он нужен для плавучести. Ростр - противовес. У некоторых мезозойских белемнитов происходила редукция ростра, тогда как аппарат для плавания (фрагмокон) - сохранялся. Так что редукция фрагмокона и ростра - процессы не всегда связанные. Эти белемниты, как считается, вели в целом пассивный образ жизни, зависая головой вниз (т.е. служили прообразом современной спирулы ) и иногда бросаясь в сторону.
Quote (heyfetzed) а) отсутствие явных рудиментов внутреннераковинных придатков у спирулы при ее активном образе жизни;
про слой - рудимент ростра я писал выше.
Активный образ жизни - спорен. Я не изучаю современных спирул, но склонен думать, что помимо Брууна спирул содержали в аквариумах и наблюдали бессчетное количество раз. И явный косяк не мог пройти незамеченным мимо десятков (если не сотен-тысяч) ученых. В этом я склонен им доверять.Quote (heyfetzed) б) сходство спирулы с наружнораковинными организмами, вкл. размеры раковины
не аргумент. Размеры - самый эволюционно изменчивый признак
Quote (heyfetzed) выступающие за контуры тела
у белемнитов, кстати, ростр тоже был обтянут тонкой прозрачной кожицей. Мы это обсуждали здесь, на форуме. Не катит
Quote (heyfetzed) пассивную защитную реакцию
функция - это не признак. Тем более, такая абстрактная, как "защита".
Quote (heyfetzed) 7. Обратная ориентация спирали у спирулы, равно как и обратное соотношение сторон у бактритоидей и белемноморф вызвано смещением ценра плавучести раковины книзу с последующим ее переворотом.
здесь обсуждать нечего, это уже ваш вывод
всё о серпулидах & белемнитах
|
| |
| |
| heyfetzed | Дата: Суббота, 23.01.2010, 17:35 | Сообщение # 33 |
|
Группа: Проверенные
Сообщений: 21
Репутация: 0
Статус: Offline
| Прикрепляю фотографию птихоцераса, найденную на сайте geo-alp.com. Отклики по основной теме последуют далее.

Сообщение отредактировал heyfetzed - Суббота, 23.01.2010, 18:02 |
| |
| |
| heyfetzed | Дата: Суббота, 23.01.2010, 17:48 | Сообщение # 34 |
|
Группа: Проверенные
Сообщений: 21
Репутация: 0
Статус: Offline
| Уважаемый Алексей Павлович,
Прежде всего, отвечу на Вашу не самую первостепенную реплику, затем перейду к другим вопросам.
На мой личный запрос рецензент (а им был К. Н. Несис) ответил, что, насколько ему известно, после Брууна спирулу в аквариуме никто не держал.
К тому же, сотни исследователей могли закрывать глаза на очевидное (фотография битой раковины тому пример), а сотням другим могли не верить либо искажать их слова. Факт, что в качестве свидетеля пассивного образа жизни спирулы ссылаются на Брууна, написавшего явно противоположное!
Сообщение отредактировал heyfetzed - Суббота, 23.01.2010, 18:08 |
| |
| |
| heyfetzed | Дата: Суббота, 23.01.2010, 18:33 | Сообщение # 35 |
|
Группа: Проверенные
Сообщений: 21
Репутация: 0
Статус: Offline
| Quote — Здесь уже обсуждать нечего. Это ваш вывод.
По-Вашему, самостоятельный вывод не может быть достойным обсуждения, тем более, если он ключевой в работе оппонента?
Напомню, что вместо того, чтобы затушевать «слабый пункт», я выявил обратную ориентацию раковины у всех колеоидей по сравнению с наружнораковинными головоногими, и рассмотрел физический механизм реориентации. Один из моих оппонентов заметил: «Это физика!» Как будто моллюски бесплотны!
Вывод является одним из объектов природы. Рецензируя статью или оппонируя на форуме, Вы должны исследовать этот объект, и выявить, является ли его связь с рассматриваемым явлением генетической, конвергентной или мимикрической. Неспособность разобрать самостоятельный вывод подразумевает неспособность увидеть новые особенности организма, который, в отличие от собеседника, не излагает их членораздельным языком.
|
| |
| |
| AlexIpp | Дата: Суббота, 23.01.2010, 19:20 | Сообщение # 36 |
|
Группа: Администраторы
Сообщений: 579
Репутация: 0
Статус: Offline
| Quote (heyfetzed) Уважаемый Алексей Павлович,
Прежде всего, отвечу на Вашу не самую первостепенную реплику, затем перейду к другим вопросам.
На мой личный запрос рецензент (а им был К. Н. Несис) ответил, что, насколько ему известно, после Брууна спирулу в аквариуме никто не держал
Это интересно: Несис действительно крупнейший ученый по современным головоногим и знал, что говорил. Но даже если по поводу образа жизни люди конкретно ошибаются, это никак не влияет на филогенетические гипотезы. Ни классические, ни вашу. Quote (heyfetzed) 7. Обратная ориентация спирали у спирулы, равно как и обратное соотношение сторон у бактритоидей и белемноморф вызвано смещением центра плавучести раковины книзу с последующим ее переворотом. Quote (heyfetzed) По-Вашему, самостоятельный вывод не может быть достойным обсуждения, тем более, если он ключевой в работе оппонента? Quote (heyfetzed) Неспособность разобрать самостоятельный вывод
мне этот вывод не кажется достаточно обоснованным. Поэтому обсуждать его я не вижу смысла. По рассуждениям, которые привели вас к нему, комментарии я дал. Мало? Quote (heyfetzed) Вывод является одним из объектов природы. Рецензируя статью или оппонируя на форуме, Вы должны исследовать этот объект
Я вам ничего не должен, и лишь даю (а точнее, безуспешно пытаюсь донести ) некоторые важные идеи по содержанию. Терпеливо. В частности, показывая, на что стоит обратить внимание при изучении данного вопроса и что прошло мимо вашего внимания. И раз на то пошло, то четких ответов на поставленные действительно важные вопросы и замечания с вашей стороны я не получил. Резюмирую это вкратце: 1. Замечание: ископаемые спирулиды не редкость, филогения на самом деле эмпирическая. Столько-то видов. Вы не учитываете стратиграфические данные. – Ответ: раковины спирул хрупкие и все равно (теоретически!) сохраниться не могли. – Хотя вам дали ссылки и даже картинку конкретной находки.
2. Замечание: Предлагая свою концепцию чего-то, не забудьте вписать туда факты, полученные другими исследователями, в частности, по мезозою – Ответ: я этого не изучал, но оно не надо
3. Какие апоморфии вы знаете у спирулид, объединяющих их с аммонитами? - ответа (четкого и однозначного) нет
4. Какие вообще факты свидетельствуют о происхождения спирулы от аммонитов? – ответа нет. Может, кто-нибудь из современных специалистов и согласится с вами обсуждать ваш вывод изолированно от рассуждений. Зеев Леви, например - он любит подобные смелые идеи. Но что-то мне подсказывает, что до сих пор большинство моих коллег либо уходили от обсуждения, либо перенаправляли вас своим знакомым. Если вы уверены в своей идее, предлагаю вам попробовать опубликовать статью в хорошем западном журнале. Если идея окажется хорошо обоснованная, ее сразу подхватят.
А я - не готов обсуждать вывод как объект природы
По крайней мере, пока не увижу ответа хотя бы на вопрос № 4. Ибо кому нужно теоретизирование, которое не подтверждается фактами? Факты нам в студию! 
всё о серпулидах & белемнитах
|
| |
| |
| mhorn | Дата: Суббота, 23.01.2010, 19:53 | Сообщение # 37 |
|
Admin
Группа: Администраторы
Сообщений: 3615
Репутация: 0
Статус: Offline
| Quote (AlexIpp) Если вы уверены в своей идее, предлагаю вам попробовать опубликовать статью в хорошем западном журнале. Если идея окажется хорошо обоснованная, ее сразу подхватят.
впрочем, публикация даже в хорошем журнале совершенно не является гарантией истинности и даже качества гипотезы. Во всяком статья уже упомянутого Зеева Леви с бредовой гипотезой происхождения аргонавтов от аммонитов была опубликована в одном из ведущих журналов Geology
верхнеюрские и нижнемеловые аммониты и всё что с ними связано
|
| |
| |
| mhorn | Дата: Суббота, 23.01.2010, 20:04 | Сообщение # 38 |
|
Admin
Группа: Администраторы
Сообщений: 3615
Репутация: 0
Статус: Offline
| PS. Про птихоцераса - вот пдф работы Догужаевой и Мутвея: http://rogov.zwz.ru/Doguzhaeva,Mutvei,1989_Ptychoceras.pdf
верхнеюрские и нижнемеловые аммониты и всё что с ними связано
|
| |
| |
| AlexIpp | Дата: Суббота, 23.01.2010, 20:04 | Сообщение # 39 |
|
Группа: Администраторы
Сообщений: 579
Репутация: 0
Статус: Offline
| Quote (mhorn) впрочем, публикация даже в хорошем журнале совершенно не является гарантией истинности и даже качества гипотезы. Во всяком статья уже упомянутого Зеева Леви с бредовой гипотезой происхождения аргонавтов от аммонитов была опубликована в одном из ведущих журналов Geology
и все же, судя по названию, имею смелость предположить, что у журнала "Geology" нет или мало рецензентов-палеонтологов И статья Леви совершенно неслучайно оказалась в итоге именно там
всё о серпулидах & белемнитах
|
| |
| |
| heyfetzed | Дата: Среда, 03.02.2010, 19:44 | Сообщение # 40 |
|
Группа: Проверенные
Сообщений: 21
Репутация: 0
Статус: Offline
| Уважаемый Алексей Павлович,
Прошу прощения за задержку из-за неполадок с интернетом!
Продолжу отвечать на замечания, с которыми успел ознакомиться в субботу 23 января.
Вначале остановлюсь на второстепенных моментах: Вы пишите: Quote «по моим представлениям, Крымгольц был одним из последних, кто выводил спирулид от белемнитов»
Впрочем, для данной работы этот вопрос третьестепенен, поскольку в главном она не согласна с обоими версиями происхождения спирулы от прямораковинных колеоидей путем спирализации — поэтому Ваше замечание («зато она /раковина спирулы/ закручена в том же направлении, что начинают закручиваться фрагмоконы ростровых спирулид»), предполагающее согласие оппонента, не учитывает действительной его позиции. Quote исследования Бизикова, на которые уже ссылался выше mhorn, однозначно показали, что на контактах внутренней раковины и мягкого тела всегда у всех групп головоногих образуется хрящ. А не минеральные образования.
Данный довод противоречит не только моим, но и Вашим соображениям, а заодно - и фактам. Отсюда следовало бы, что ни одна мягкая часть тела внутреннераковинных головоногих неспособна выработать минеральный рострум на контакте с внутренней раковиной. Как же тогда он возникает? Естественно, что его вырабатывает часть мантийного мешка прилегающая к наружной стенке фрагмокона. Полагаю, что Бизиков писал про хрящ, но не против минералов.
В данном случае основной предмет спора состоит в утолщении стенки раковины спирулы, который Вы принимаете за рудимент рострума. Если это не иллюзия (на своем экземпляре я его не увидел; через стенки раковины везде просвечивают септы), то такое утолщение столь слабо, что должно быть названо следом либо предпосылкой, а не рудиментом или зачатком (и тот, и другой по форме напоминают развитый орган). Я утверждаю, что это могла быть предпосылка возникновения рострума. Понятно, что ростровые формы произошли от каких бы то ни было безростровых. Если же принять редукцию придатков спирулы более глубокой, чем в значительно сильнее модифицированном сепионе (кстати, Вы считаете, что спирулинять версию а возникла в миоцене, тогда как каракатицы известны с юры), — следует предположить, что ничтожная тяжесть значительно снижала шанс спирул на выживание… В чем же ее значение? Добро, если бы спирула жила в Черном море с сероводородной зоной, или в Байкале при дефиците солей! В океане же соли хватает, да и по вертикали спирула мигрирует в весьма широких пределах — от сотен метров до километров. Не соглашаясь со мной в пункте редукции фрагмокона у потомков спирулы, Вы пишите:
«В общем, если описывать кратко, то ситуация понимается учеными так», а затем приводите общепринятую версию эволюции спирулид. Но общепринятость не есть аргумент — поколения астрономов придерживались концепции Аристотеля и Птолемея о неподвижности Земли. Чье понимание вернее, решает сопоставление с природой. В противном случае происходит не поиск истины, а игра в науку по определенным правилам. Quote предлагаю для сравнения посмотреть на картинку. Этот вопрос (радиус обзора) - классика...
Комментировать ничего не буду.
Предлагаю удостовериться в соответствии приведенной Вами иллюстрации с моими словами. Повторю их: расстояние м. глазами спирулы, следовательно, диаметр окружности, проведенной через них, гораздо меньше продольной оси тела.
Недаром Несис, исходя из существующей версии, объявляет функцию светящегося органа таинственной. По поводу пассивной защиты спирулы, сходной с наружнораковинными моллюсками, Вы пишите: функция — это не признак. Тем более, такая абстрактная, как "защита".
Между тем, для животного вопрос жизни и смерти отнюдь не абстрактен. Ученый действительно должен владеть абстрактным мышлением для сопоставления статических признаков и функций, как это делал, к примеру, великий Кювье. Теперь о главном:
Базисным положением работы, из которого вытекают все прочие следствия, и которое я постарался обосновать дополнительно, является неадаптивность спирализации фрагмокона для увеличения плавучести.
Вы предлагаете дополнительные резоны спирализации:
Quote Спиральный фрагмокон дает возможность сместить центр плавучести вперед.
Спирализация даже у ростровых форм несколько увеличивает расстояние между центром плавучести и задним центром тяжести. Как результат, на построение противовеса идет меньшее количество материала, что выгоднее энергетически.
Уже зависание спирулы вниз головой в состоянии покоя свидетельствует не в пользу смещения плавучести вперед (т. е. к голове). Что касается вымерших белемнозеин, то изгиб фрагмокона находится в задней части. Поэтому его объем, а значит, и плавучесть увеличиваются здесь, а не спереди.
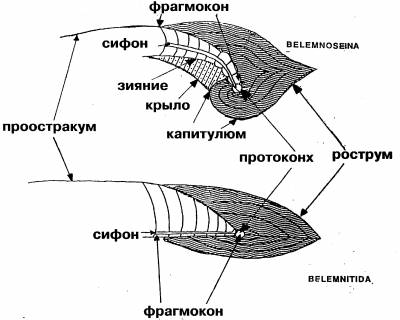
Соответственно, кзади (а не кпереди, как Вы утверждаете) смещен и центр плавучести у более изогнутых белемнозеин в сравнении с менее изогнутыми. Таким образом, спирализация фрагмокона при сохранении массивного рострума неадаптивна, поскольку:
• Уменьшает полезный объем тела (который мог бы высвободиться редукцией рострума).
• Увеличивает расход материала на строительство фрагмокона (тогда как редукция рострума уменьшила бы его).
• Приближает центр плавучести фрагмокона к центру тяжести рострума. Т. о. спирализация фрагмокона противодействует функции рострума.
Поскольку естественный отбор лишь сохраняет благоприятные изменения, более изменчивая из эволюционирующих частей (в данном случае, рострум), скорее отображает тенденции эволюции. В пользу этого свидетельствует и приведенный Вами пример безростровых белемнитов. Сохранение массивного рострума в различных группах подотряда белемнозеин на фоне различной степени изгиба относительно консервативного фрагмокона свидетельствуют о редукции не первого, а последнего; приспособлении не к увеличению плавучести, а к ее уменьшению, позволяющему уравновесить тело в горизонтальном положении.
Ваш вывод следует из сопоставления спирулы и форм с массивным рострумом, но не подтверждается «промежуточными стадиями». Как я уже писал, рострум спирулиростры развит не хуже, чем у белемнозиса. N. B. Поначалу не вчитался в Ваш текст. Вы пишете: Quote «И явный косяк не мог пройти незамеченым мимо десятков (если не сотен тысяч) ученых. В этом я склонен им доверять»
С точки зрения принятой версии спирула обязана быть пассивной. Описание Брууна, вроде бы, противоречит хорошо обоснованной теоретической идее. С другой стороны, не вызывает сомнения и основательность экспедиции. Здесь на помощь приходит способность к интерпретации. Слова о том, что спирула повисла вниз головой, бросаются в глаза, тогда как на последующие просто не обращают внимания, не вчитываются — Вы сами показали пример этого. Скажу больше: ознакомившись с отчетом Брууна, поначалу не заметил этих строк и я, поскольку придерживался общепринятого взгляда. Впоследствии, придя к выводу об активном образе жизни спирулы, успел написать, якобы условия ее содержания в аквариуме способствовали неверному выводу, и лишь, обратившись к отчету повторно, прочел предельно ясный текст. По поводу дополнительных наблюдений:
Допустим, помимо экспедиции Брууна спирулу наблюдали в аквариуме представители иной экспедиции, и нашли ее пассивной.
Допустим, что непосредственные наблюдения в обоих случаях велись добросовестно. Что легче: ввергнуть активное глубоководное животное при подъеме на поверхность в состояние глубокого шока или заставить пассивное животное с деградировавшей мускулатурой не судорожно дергаться, а превосходно плавать (даже раковиной вниз) в ограниченном объеме аквариума в течение 48 ч?
Если спирула является активным организмом, если она преодолевает отклоняющую силу плавучести раковины собственными усилиями, то отсутствие рострума у нее не вторично, а первично. Предлагаю удостовериться в подлинности строк отчета Брууна.
Подвожу итог сказанному:
• Спирула является активным организмом, происходящим от спиральных форм лишенных рострума.
• Спирализация внутренней раковины неадаптивна по причинам, указанным выше.
• Спирула происходит от наружнораковинных аммонитов.
• Реориентация ее раковины, а также раковины прямых колеоидей в сравнении с наружнораковинными головоногими объясняется изменением распределения сил плавучести с дальнейшим внезапным переворотом.
Предлагаю признать обоснованность работы и перенести дискуссию на страницы академических журналов. Готов внести в текст соответствующие изменения.
Сообщение отредактировал heyfetzed - Среда, 03.02.2010, 19:52 |
| |
| |
| heyfetzed | Дата: Среда, 03.02.2010, 19:49 | Сообщение # 41 |
|
Группа: Проверенные
Сообщений: 21
Репутация: 0
Статус: Offline
| Вы пишете: Quote мне этот вывод не кажется обоснованным
Я уже писал о позиции великого Кювье в отношении эхолокации у летучих мышей. Обоснованность или необоснованность вывода проверяется доводом, а не авторитетом.
Сообщение отредактировал heyfetzed - Среда, 03.02.2010, 19:58 |
| |
| |
| AlexIpp | Дата: Четверг, 04.02.2010, 00:39 | Сообщение # 42 |
|
Группа: Администраторы
Сообщений: 579
Репутация: 0
Статус: Offline
| Вы удивительный человек
Я желаю вам успешно отстаивать вашу гипотезу и дальше, на страницах академических журналов.
всё о серпулидах & белемнитах
|
| |
| |
| mhorn | Дата: Четверг, 18.02.2010, 18:25 | Сообщение # 43 |
|
Admin
Группа: Администраторы
Сообщений: 3615
Репутация: 0
Статус: Offline
| любителям спирул - свежая статья:
Warnke K.M., Boletzky S.v. The ”ventral rib” of the shell in Spirula (Cephalopoda, Coleoidea): a cryptic rostrum? // Berliner paläobiologische abhandlungen. Bd.10. P.357-359.
Pdf: http://www.geo.fu-berlin.de/geol....zky.pdf
После более чем столетнего перерыва переоткрыта мембрана, которая соединяет у спирулы начальную камеру и вентральную поверхность последующих камер. Эта мембрана может служить в качестве направляющей и укрепляющей структуры во время образования первых камер. Её эволюционное происхождение, видимо, связано с ростром.
верхнеюрские и нижнемеловые аммониты и всё что с ними связано
|
| |
| |
| AlexIpp | Дата: Воскресенье, 14.03.2010, 20:52 | Сообщение # 44 |
|
Группа: Администраторы
Сообщений: 579
Репутация: 0
Статус: Offline
| вчера получил от Эдуарда Хейфеца PDF замечательной работы про спирулу: Bruun, A.F. 1943. The biology of Spirula spirula (L.) // Dana-Report. No.24. 49pp.
http://rogov.zwz.ru/bruun,%....ula.pdf
всё о серпулидах & белемнитах
|
| |
| |
| heyfetzed | Дата: Пятница, 24.12.2010, 11:46 | Сообщение # 45 |
|
Группа: Проверенные
Сообщений: 21
Репутация: 0
Статус: Offline
| Сообщением о статье Варнке и Болетцки предполагаю воспользоваться. Авторы углядели рострум на внутренней стороне спирали, т. е. там, где он отсутствует у ближайших родственников спирулы. Логичнее предположить, что это след контакта оборотов у плотноспиральной предковой формы.
Сообщение отредактировал heyfetzed - Пятница, 24.12.2010, 11:47 |
| |
| |
| mhorn | Дата: Суббота, 25.12.2010, 10:37 | Сообщение # 46 |
|
Admin
Группа: Администраторы
Сообщений: 3615
Репутация: 0
Статус: Offline
| Quote (heyfetzed) Авторы углядели рострум на внутренней стороне спирали, т. е. там, где он отсутствует у ближайших родственников спирулы.
почему отсутствует?
Quote (heyfetzed) Логичнее предположить, что это след контакта оборотов у плотноспиральной предковой формы.
почему? у тех же гетероморфных аммонитов подобные образования отсутствуют
верхнеюрские и нижнемеловые аммониты и всё что с ними связано
|
| |
| |
| heyfetzed | Дата: Воскресенье, 26.12.2010, 13:15 | Сообщение # 47 |
|
Группа: Проверенные
Сообщений: 21
Репутация: 0
Статус: Offline
| У ближайших родственников спирулы рострум развивается на дорсальной, а не на вентральной стороне. Данная мембрана отсуствует как у гетероморфных аммонитов, так и у родственников спирулы - она слишком хрупка (недаром ее так долго не замечали), чтобы сохраняться в ископаемом состоянии.
|
| |
| |
| Ecolog | Дата: Понедельник, 10.10.2011, 10:58 | Сообщение # 48 |

Группа: Проверенные
Сообщений: 123
Репутация: 0
Статус: Offline
| В этой дискуссии оказались затронуты очень интересные вопросы. с которыми хотелось бы разобраться.
Quote (AlexIpp) мантия и мускулистая мантия - это разные вещи. Разбирайтесь. Мускулистая мантия окружает полукольцом все мягкое тело и сверху крепится к краям проостракума (пока он был). У ростровых спирулид - частично и к боковым краям ростра.
У ростровых спирулид, получается, должны были оставаться отпечатки мускулов? А где про это можно прочитать? И у спирулы - сейчас, когда нет проостракума - мускулистая мантия к чему-нибудь крепится?
Quote (mhorn) втягивать голову в мантию могут многие головоногие
А как спирула это делает? То есть у наутилуса мощные мускулы-ретракторы крепятся к раковине, у кальмаров, которые могут втягивать голову в мантию ретракторы крепятся к гладиусу. А у спирулы они к чему крепятся, к этой раковине? Или там вообще как-то по-другому все организовано? И опять же, где можно про это почитать?
UPD. часть ответов нашел в книге "FOSSI DIBRANCHIATE CEPHALOPODS" A.Naef, 1922, но буду очень благодарен за ссылки на другие источники. Должны же быть и современные публикации о мускулатуре вымерших и живых спирулид.
Любитель аммонитов и прочих головоножек
Сообщение отредактировал Ecolog - Понедельник, 10.10.2011, 11:27 |
| |
| |
|
 Форум Jurassic.ru
Форум Jurassic.ru